Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника

Этапы пути инициации репликации на ОНР у дрожжей
|
|
В конце митоза или в начале фазы G1 клеточного цикла нуклеопротеиновые комплексы ORC-ARS вербуют на ДНК белок Cdc6 c мол. массой 58 кД. Этот белок очень нестабилен и должен синтезироваться de novo после выхода клеток из митоза. Он имеет несколько доменов гомологии с субъединицами Orc1, Orc4 и Orc5 комплекса ORC и взаимодействует с ним. Кроме того, Cdc6 гомологичен белкам g-комплекса бактериальной ДНК-полимеразы III и эукариотического комплекса RFC – многосубъединичным ферментам, катализирующим зависящую от гидролиза АТФ погрузку на ДНК скользящих зажимов ДНК-полимераз. Подобно этим ферментам класса АТФаз ААА+, белок Cdc6 способен связывать и медленно гидролизовать АТФ in vitro. Консервативный домен связывания нуклеотидов в Cdc6 необходим для функционального взаимодействия с ORC-ARS in vivo. По аналогии с погрузчиками факторов процессивности, считается, что Cdc6 является погрузчиком на хроматин кольцевого комплекса MCM – ДНК-геликазы репликативных вилок, концентрация которой в клетках дрожжей в 10-100 раз больше концентрации ORC. Белок Cdc6 преимущественно взаимодействует в ORC с комплексом Orc1-АТФ. После узнавания ORC белком Cdc6, связавшим АТФ, образуется комплекс ARS-ORC-Cdc6, который, используя катализируемый Cdc6 гидролиз АТФ, привлекает к ОНР белки Mcm2-Mcm7. В этом процессе у S. cerevisiae участвует также не гомологичный субъединицам гексамера МСМ белок Mcm10, взаимодействующий с Mcm7. Отметим также, что связанный с ORC белок Cdc6 cпособствует ассоциации фактора транскрипции Abf1 c элементом В3 в ARS.
Погрузка геликазы МСМ на ORC-ARS в начале фазы G1 завершает образование “ предрепликативного комплекса ”, в котором ОНР получила “ лицензию на репликацию ” и перешла в компенентное для инициации состояние. Однако “запуск” (firing) репликации на уже готовой к инициации ОНР откладывается до фазы S клеточного цикла. Для такого запуска необходимо действие циклин-зависимых протеинкиназ, которые появляются только в начале этой фазы. К ним относятся комплексы главной киназы Cdc28 c циклинами типа В (Clb5 и Clb6) и киназы Cdc7 c её регуляторным циклиноподобным белком Dbf4. Циклины Clb5 и Clb6 синтезируются уже в фазе G1 и ассоциируются с Cdc28, но эти комплексы остаются неактивными до начала фазы S, когда их ингибитор Sic1 фосфорилируется под действием комплексов Cdc28 с циклинами фазы G1 и подвергается зависящему от убиквитина протеолизу. Белок Dbf4 очень нестабилен в течение всего клеточного цикла, особенно в начале фазы G1, когда его период полураспада равен 5 мин. Однако в начале фазы S его уровень и активность протеинкиназы Cdc7-Dbf4 достигают максимума. Таким образом, оба типа протеинкиназ становятся активными почти одновременно. Однако в активации ими ОНР проявляется определенная иерархия: Cdc7-Dbf4 действует после Cdc28-Clb.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Одним из субстратов для Cdc28-Clb является белок Cdc6, который сыграл свою роль после погрузки комплекса МСМ. Этот белок после фосфорилирования покидает комплекс с ORC и подвергается протеолитической деградации. В освобождении связанного Cdc6 может играть роль гидролиз АТФ, связанного с белком Orc1. На освободившееся место вербуется новый важный компонент инициации репликации – белок Cdc45, который взаимодействует с белками МСМ. Для включения Cdc45 в предрепликативный комплекс требуется действие киназы Cdc7-Dbf4, которая связывается с белками МСМ и вызывает их фосфорилирование. Белки Mcm2-4 и Mcm6-7 являются субстратами для этой киназы in vitro. Фосфорилирование МСМ под действием Cdc7-Dbf4 сопровождается повышением чувствительности ДНК в области В2 ARS1 к KMnO4, взаимодействующему с онДНК. Это показало, что Cdc7-Dbf4 запускает переход ДНК геликазы МСМ в активное состояние (см. 2.1) и вызывает локальное расплетание ДНК в богатом А: Т сегменте ОНР. Связывание Cdc45 и модификация MCM приводят к образованию “ преинициирующего комплекса ”. В этот комплекс привлекаются также связывающий онДНК белок RPA и ДНК-полимераза a - праймаза. Вербовка этих белков критически зависит от присутствия в преинициирующем комплексе белка Cdc45, который может физически взаимодействовать с ними. В частности, белок Cdc45 связывается с субъединицей р70 комплекса Pola-праймаза (рис. 00). Киназа Cdc7-Dbf4 способна фосфорилировать и такие компоненты репликативного комплекса, как ДНК-полимераза a. Вербовка Pola-праймазы является первым этапом образования репликативных вилок на ARS.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
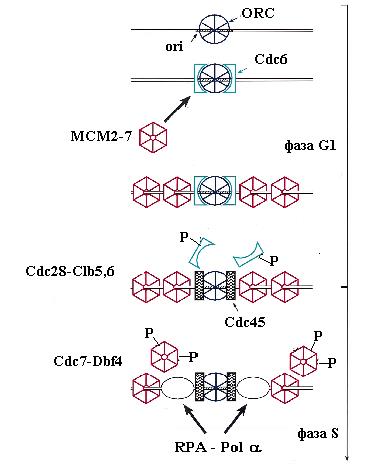
Рис. 3.7. Этапы пути инициации репликации ДНК у дрожжей
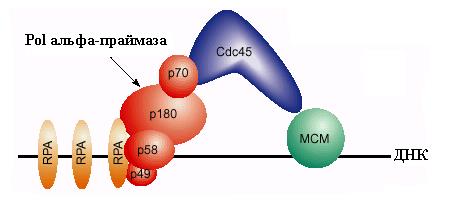
Рис. 3.8. Схема участия белков МСМ, ДНК-полимеразы a - праймазы и белков Cdc45 и
RPA в инициации репликации на эукариотической области ori
После образования двунаправленных репликативных вилок геликаза МСМ и белок Cdc45 выходят из контакта с комплексом ORC и перемещаются вместе с репликативной ДНК-полимеразой. На ДНК ARS остается только пострепликативный комплекс, содержащий ORC – как и в начале пути инициации репликации. Такие комплексы существуют в течение митоза и ранней фазы G1 на копиях ARS в обеих дочерних хромосомах.
|
|