Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Для новых пользователей первый месяц бесплатно. Чат-бот для мастеров и специалистов, который упрощает ведение записей: — Сам записывает клиентов и напоминает им о визите;
— Персонализирует скидки, чаевые, кэшбэк и предоплаты;
— Увеличивает доходимость и помогает больше зарабатывать; Начать пользоваться сервисом

I II III
|
|
B.



 1 115 165 177
1 115 165 177
I II III
C.
1 181 290 301 422 432 616



 RPA1 DBD-A DBD-B DBD-C
RPA1 DBD-A DBD-B DBD-C

 Pola
Pola
 p53
p53
 RPA2
RPA2
 1 43 171 270
1 43 171 270
 RPA2 DBD-D
RPA2 DBD-D
 RPA1
RPA1

RPA3
1 121
Рис. 2.12. Домены связывающих онДНК белков.
А. Белок gp32 фага Т4.
I – домен, ответственный за кооперативность связывания с онДНК, II – минимальный домен связывания с онДНК, III – домен взаимодействия с другими белками.
В. Белок SSB E. coli.
I – домен связывания с онДНК, олигомеризации и кооперативности, II – домен статистического клубка, участвующий во белок-белковых взаимодействиях, III – участок гомологии с кислой областью на С-конце gp32.
C. Эукариотический белок RPA.
Указаны положения сайтов связывания с онДНК (DBD) субъединицах RPA1 и RPA2 и контактных участков белок-белковых взаимодействий
Белок SSB E. coli кодируется существенным геном, расположенным на 92-ой мин генетической карты и имеющим SOS-индуцибельный промотор. Температурочувствительные мутации ssb летальны при непермиссивной температуре, при которой они останавливают репликацию ДНК. Кроме того, мутации повышают чувствительность клеток к повреждающим ДНК агентам и вызывают аномалии рекомбинации и дефект по индукции SOS-системы. Физиологически активной формой белка SSB является тетрамер, число копий которого на клетку равно 300-350. Этого количества достаточно, чтобы покрыть белком SSB участок длиной 1400 н. на репликативную вилку. Белок SSB имеет длину 177 остатков и мол. массу 18, 9 кД и состоит из 3 доменов (рис. 2.12, В). Наиболее важной является N-концевая область (остатки 1-105), являющаяся доменом связывания с ДНК. Она содержит также аминокислотные остатки, существенные для взаимодействия мономер-мономер в тетрамере SSB и для кооперативности связывания SSB с онДНК. Домен II, состоящий из остатков 106-165, находится в конформации статистического клубка и не содержит заряженных остатков. После связывания SSB с онДНК этот домен становится более чувствительным к протеолизу. Это показало, что домен II доступен для взаимодействия с другими белками, и осуществляет функции белок-белковых взаимодействий. Последние 12 остатков на С-конце белка SSB очень похожи на кислую С-концевую область белка gp32. У обоих белков протеолитическое удаление С-концевых остатков изменяет кинетику связывания с ДНК и повышает способность дестабилизировать днДНК. Эта область SSB также важна для взаимодействия с другими белками, участвующими в метаболизме ДНК.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Эукариотический белок RPA является гетеротримерным и состоит из 3 разных субъединиц: RPA1, RPA2 и RPA3. Гены этих белков у человека расположены соответственно в хромосомах 17, 1 и 7, а сами субъединицы имеют длину 616, 270 и 121 аминокислотных остатков (мол. массы» 70, 29 и 14 кД). Эти белки гомологичны белкам Rfa1, Rfa2 и Rfa3 субъединиц гетеротримерного фактора RFA S. cerevisiae. У дрожжей гены всех 3 субъединиц связывающего онДНК белка абсолютно необходимы для жизнеспособности. Все субъединицы человеческого белка RPA требуются для репликации ДНК вируса SV40 in vitro.
Cубъединица RPA1 (рис. 2.12, С) может связываться с онРНК самостоятельно, но этого недостаточно для поддержания репликации ДНК. Эта субъединица является главной в гетеротримерном комплексе RPA. Её N-концевой домен (NTD) является наиболее важным участком во взаимодействиях с другими белками, хотя некоторые из этих белков имеют дополнительный сайт взаимодействия и на С-конце RPA1. RPA1 связывается с такими белками систем репликации ДНК, как большой Т-антиген вируса SV40, узнающий вирусную область инициации репликации, и с ДНК-полимеразой a - праймазой. Во взаимодействиях с этими белками участвуют N- и С-концевые области RPA1. Кроме того, RPA1 ассоциируется с активаторами транскрипции, имеющими кислые домены активации (например, с белком VP16 вируса простого герпеса и клеточным белком р53), а также с белком XP-G репарации ДНК. Домен взаимодействия с р53 расположен только в N-концевой области RPA1. Эти взаимодействия могут играть роль в привлечении RPA к сайтам репликации и репарации.
Субъединица RPA1 содержит также 3 из 4 доменов связывания RPA с онДНК: DBD-A (остатки 181-290), DBD-B (остатки 300-422) и DBD-C (остатки от 432 до ~560). Между этими доменами расположены гибкие линкерные области. При связывании с онДНК тандема доменов DBD-А и DBD-В каждый из них контактирует с 3 н., а промежуточные 2 н. онДНК приходятся на междоменную область. Четвертый слабый участок связывания с онДНК (домен DBD-D) расположен в центре субъединицы RPA2 и занимает более пРоловины этой молекулы (остатки 43-171). Самая маленькая субъединица RPA3 прямо не связывается с онДНК и её функции неизвестны. Предполагается, что С-концевая область RPA2 служит мостиком между RPA1 и RPA3. В тримеризации RPA участвуют три С-концевые a-спирали субъединиц RPA. Тримеризация RPA наиболее существенна для взаимодействия RPA с длинными субстратами онДНК (> 24 н.), а не с короткими олигонуклеотидами (10 н.).
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
Рассмотрим молекулярные аспекты взаимодействия связывающих онДНК белков с их “субстратами” онДНК. Все белки кооперативно ассоциируются с онДНК, но возможны два типа кооперативного связывания. При “неограниченной” кооперативности белок образует на онДНК длинные кластеры, в которых каждая молекула белка взаимодействует с ближайшими соседями по обе стороны от неё. В предельном случае при параметре кооперативности w®¥ белок полностью покрывает онДНК. Для “ограниченной” кооперативности характерно образование на ДНК “бусинок” - отдельных ограниченных групп кооперативно ассоциированных белковых молекул. Различные моды кооперативного связывания могут избирательно использоваться в процессах репликации, рекомбинации и репарации. Детальные механизмы образования контактов с онДНК неодинаковы для разных типов связывающих онДНК белков. Их можно рабить на два класса.
Белок SSB E. coli, являющийся в растворе стабильным тетрамером, связывается с онДНК в форме тетрамера с константой ассоциации 108-1010 М-1. Для белка SSB возможны три моды связывания с ДНК, в которых он покрывает участки онДНК длиной 35±2, 56±3 и 65±3 н. Эти моды названы соответственно (SSB)35, (SSB)56 и (SSB)65. Для моды (SSB)35 характерна неограниченная кооперативность. В моде (SSB)63 проявляется ограниченная кооперативность, при которой “бусинки” на онДНК соответствуют “октамерам” – кооперативно взаимодействующим смежным тетрамерам SSB. Мода (SSB)65 преобладает при низкой (< 0, 01 M NaCl), а мода (SSB)35 – при высокой (> 0, 2 M NaCl) ионной силе. При физиологических значениях ионной силы существует равновесная смесь этих двух мод связывания. Уже в 80-е годы в качестве рабочей модели взаимодействия белка SSB с онДНК была предложена следующая схема (рис. 2.13). В моде (SSB)35 только две из 4 субъединиц каждого тетрамера SSB взаимодействуют с онДНК, а в модах (SSB)56 и (SSB)65 в ассоциации с онДНК участвуют все 4 субъединицы. Во всех случаях онДНК связывается с поверхностью асимметричных субъединиц SSB и наматывается на поверхность белковой сердцевины тетрамера с образованием структуры, очень похожую на нуклеосому. В моде (SSB)65 два смежных тетрамера ассоциированыы друг с другом в октамер, на который намотан участок он ДНК длиной ~145 н. На участках длиной ~30 н. между двумя соседними октамерами SSB онДНК свободна от белка. Образование нуклеосомоподобной структуры в моде (SSB)65 сопровождается уменьшением контурной длины онДНК на 75%.
] 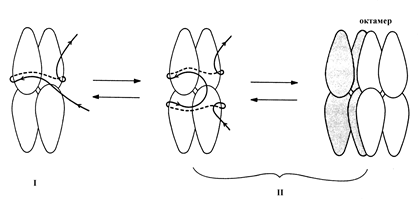
Рис. 2. 13. Модель предполагаемой организации комплексов белка
с онДНК.
I – мода связывания (SSB)35, II – моды связывания (SSB)56 и (SSB)65
Эта модель ещё не полностью доказана экспериментально. Только в 1997 г. методом рентгеностструктурного анализа с разрешением 2, 9 А удалось установить структуру минимального домена связывания с онДНК, состоящего из остатков 1-135 белка SSB E. coli. Оказалось, что мономерная форма этого домена имет очень асимметричную форму, похожую на уголок (рис. 2.14, А). По своей структуре мономер SSB относится к большой группе белков с укладкой типа ОВ (оligomer binding – связывание олигомеров), связывающихся с олигонуклеотидами и олигосахаридами. Укладка ОВ является вариантом широкораспространенной укладки “греческого ключа”. В “основании” уголка SSB расположены a-спираль из остатков 61-70 и несколько b-нитей (нити 1-5). Из этого основания выступает сильно вытянутая петля L45 остатков 82-101, образующая b-шпильку 451-452.
В тетрамерах белка SSB объединяются b-нити оснований из всех 4 субъединиц SSB с образованием 6-нитевых b-плоскостей. Шпильки петель L45 смежных димеров, расположенных над или под плоскостью, попарно взаимодействуют друг с другом. В результате тетрамер SSB приобретает симметричную форму, похожую на эллипсоид вращения (рис. 2.14, В). На поверхности тетрамера расположены ароматические остатки трп54, трп88 и фен60, участвующие в стэкинг-взаимодействиях с основаниями онДНК, а также положительно заряженные остатки лиз62 и лиз73, важные для связывания с остовом онДНК (рис. 2.14, С).Взаимодействие с остатками, поставляемыми каждым из мономеров тетрамера SSB и переход от одного мономера к другому, вероятно, определяют геометрию наматывания онДНК на поверхность тетрамера. Однако в комплексах типа (SSB)56 и (SSB)65 она пока не установлена. Если в комплексах с SSB онДНК упаковывается на поверхности, то в комплексах с фаговым белком gp32 и эукариотическим белком RPA она, скорее, размещается во впадинах или каналах.
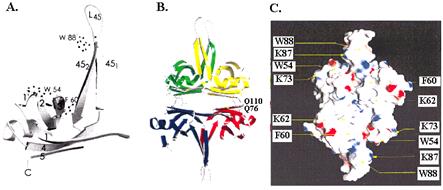 |
Рис. 2.14. Структура минимального домена связывания с онДНК (остатки 1-135) белка ` SSB E. coli.
A – мономерная форма, В – тетрамерная форма, С - распределение аминокислотных остатков, важных для связывания онДНК, на поверхности белка
Активной формой белка gp32 фага Т4 является мономер, который связывает участок онДНК длиной 6-7 н. Ассоциация gp32 с онДНК происходит с очень высокой, но не бесконечно большой кооперативностью, так что этот белок образует очень длинные кластеры на онДНК, но не покрывает её полностью. Связывание gp32 увеличивает контурную длину онДНК. Рентгеноструктурный анализ (разрешение 2, 2 А) комплекса тетрануклеотида онДНК с минимальным связывающим онДНК доменом gp32 (остатки 22-239) показал, что этот домен содержит пять a-спиралей и восемь b-нитей. Домен состоит из трех субструктур: нижнего субдомена I, координационно связывающего Zn2+ с одним остатком гистидина и 3 остатками цистеина, верхнего субдомена II, имеющего структуру 5-нитевого b-слоя, и соединяющей их области из одной a-спирали и двух b-нитей. В домене связывания онДНК имеется положительно заряженная центральная «щель» с гидрофобными карманами для связывания оснований ДНК. Карманы содержат существенные ароматические аминокислотные остатки тир и фен, поставляемые b-нитями субдоменов I и II. Эта полость наиболее прочно связывает участок онДНК длиной 2-3 н. Связанная онДНК находится в растянутой конформации, в которой расстояние между соседними основаниями равно 5 Å, а не 3, 4 Å, как в стандартной В-форме ДНК.
Очень похожий механизм ассоциации с онДНК обнаружен и для эукариотического белка RPA. Хотя самая большая субъединица RPA1 может самостоятельно связывать онДНК, физиологически активной является гетеротримерная форма. RPA связывается с онРНК в определенной полярности и имеет три основные моды ассоциации. В главной моде, имеющей высокое сродство к онДНК, но низкую кооперативность, тример RPA маскирует участок онДНК длиной 30 н. Вторая мода менее стабильна и, вероятно, является предшественником первой. В ней белок RPA проявляет более низкое сродство и более высокую кооперативность. Переход между этими двумя модами является физиологически важным событием для расплетания ДНК и происходит через промежуточное состояние, в котором RPA связывается с 13-14 н. в онДНК.
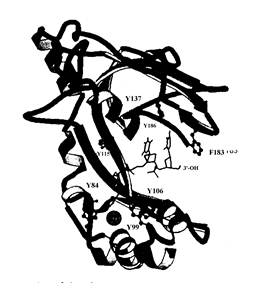
Рис. 2.15. Структура комплекса минимального домена связывания онДНК белка gp32 фага Т4.Указаны положения связанного катиона Zn2+, 3’-конца олигонуклеотида онДНК и нескольких ароматических аминокислотных остатков, существенных для связывания он ДНК
Структура минимального домена связывания с онДНК, состоящего из двух доменов (DBP-A и DBP-B) субъединицы RPA1 (остатки 181-422), и его комплекса с онДНК определена с разрешением 2, 5 Å. В свободном состоянии домены А и В, соединенные гибким линкером (остатки 291-299), имеют укладку типа ОВ, важным признаком которой является 5-нитевой b-слой, сворачивающийся в b-бочку. В отсутствие ДНК тандем домеров А и В может находиться в двух разных конформациях: менее компактной и более компактной (рис. 2.16, А и В). Тандемная ориентация укладок ОВ становится ещё более компактной и значительно стабилизируется в присутствии онДНК (рис. 2.16, С). В комплексе с DBD1-DBD2 онДНК располагается во внутреннем канале белка диаметром ~17 Å. Каналы связывания онДНК в доменах А и В ориентированы в одном направлении, так что ДНК переходит из одного домена в другой по прямой линии. В образовании этих каналов важную роль играют вытянутые петли L12, которые образуют b-шпильки, как в белке SSB, и охватывают ДНК, подобно пальцам. В связыввании онДНК участвуют стэкинг с двумя ароматическими остатками (двумя остатками фен в домене А и остатками фен и трп в домене В) и водородные связи между аминокислотными остатками белка и основаниями и фосфатными группами онДНК. Домен А образует больше контактов с онДНК, чем домен В. Каждый из этих доменов взаимодействует с 3 н. онДНК и ещё 2 н. приходятся на долю линкера между доменами. Таким образом, структура комплекса с доменами DBP-A и DBP-B полностью объясняет моду связывания RPA c участком онДНК длиной 8 н.
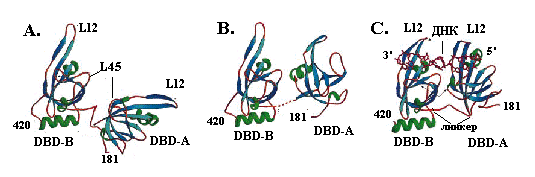
Рис. 2.16. Структура тандема доменов DBP-A и DBP-B субъединицы RPA1 белка RPA.
А и В – две альтернативные конформации доменов в отсутствие ДНК, С – комплекс белка с онДНК
Для объяснения двух других мод связывания RPA необходимо привлечь ещё два домена (DBP-С на С-конце RPA1 и DBP-D в середине RPA2), которые, предположительно, также имеют укладку типа ОВ. Сборка моды связывания с 30 н. онРНК может происходить в несколько этапов (рис. 2.17). Первый из них состоит в связывании доменов DBP-A и DBP-B с участком длиной 8 н. Этот процесс инициируется доменом А, которые имеет более высокую активность и гибкость т первым ассоциируется с 5’-концом онДНК. Затем к нему присоединяется домен DBP-B. Последовательное связывание доменов А и В объясняет известную полярность (5’®3’) ассоциации RPA c онДНК. В результате первичного связывания тандема А+В белок RPA переходит из глобулярной формы (рис. 2.17, А) в более вытянутую (рис. 2.17, В). Изменение конформации сбъединицы RPA1 дает возможность связаться с онДНК следующему домену DBP-С, которые вместе с линкером между доменами В и С также должен занимать на онДНК участок длиной 5 н. В результате белок RPA переходит в промежуточную моду связывания с 13-15 н. На заключительном этапе присоединяется домен DВР-D из второй субъединицы RPA, связывающийся с 3’-стороны от DBP-С (рис. 2.17, D). Квартет доменов А, В, С и D, каждый из которых (вместе с линкером) занимает не более 5 н. на онДНК, образует сердцевину (18-20 н.) моды связывания RPA с 30 н. онДНК. Недостающие 10-12 н. в этой моде, вероятно, приходятся на долю N-концевого домена (NTD) RPA1 и области RPA2 за пределами домена DBP-D.
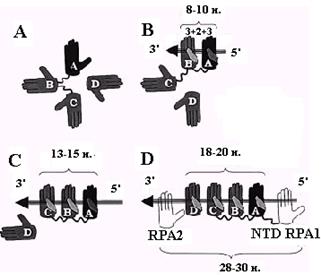
Рис. 2.17. Последовательные этапы сборки мод связывания белка RPA c участком онДНК длиной 30 н.
Домены DВР-А - DВР-D белка RPA изображены в виде ладоней, а онДНК – в виде стрелки
|
|