Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника

Бактериальные ДНК-полимеразы
|
|
Характеристики главных типов бактериальных ДНК-полимераз наиболее детально изучены на примере E. coli (табл. 1.2). Всего у кишечной палочки существуют 5 разных ДНК-полимераз. Мы рассмотрим вначале только три из них, отложив характеристику двух «склонных к ошибкам» ДНК-полимераз до гл. 00.
Таблица 1.2
ДНК-полимеразы E. coli
| ДНК-полимеразы | Ген | Положение на карте, мин | Продукт | Число остатков | Мол. масса кД | Функция |
| ДНК-полимераза I (A) | polA | 87, 2 | PolI | Репарация, репликация | ||
| ДНК-полимераза II (B) | polB | 1, 4 | PolII | Репарация, мутагенез | ||
| Главная репликативная ДНК-полимераза III (C) | dnaE | 4, 4 | a, PolIII | Каталитическая субъединица | ||
| dnaN | 83, 6 | b, PolIII | Скользящий зажим | |||
| dnaQ | 5, 1 | e, PolIII | (3’®5’)-экзонуклеазная субъединица | |||
| dnaX | 10, 6 | t, PolIII g, PolIII | Структурная субъединица | |||
| Комплекс g-комплекса погрузчика зажима | ||||||
| holA | 14, 4 | d, PolIII | “ | |||
| holB | 24, 9 | d’, PolIII | “ | |||
| holC | 96, 6 | c, PolIII | “ | |||
| holD | 99, 3 | y, PolIII | “ | |||
| holE | 41, 5 | q, PolIII | Компонент минимального фермента |
1.2.1. ДНК-полимераза I E.coli
Из всех ДНК-полимераз E. coli наиболее высокий внутриклеточный уровень (~ 400 молекул на клетку) имеет ДНК-полимераза I, кодируемая существенным геном polA. Она состоит из одной субъединицы длиной 928 остатков, которую можно разделить на три домена (рис. 1.6). N-концевую треть молекулы занимает (5’®3’)-экзонуклеазный домен, в центре расположен самый короткий (3’®5’)-экзонуклеазный домен, а самый большой полимеразный домен находится на С-конце. При ограниченном протеолизе ДНК-полимеразы I расщепление на границе между первыми доменами дает малый N-концевой 5’-нуклеазный фрагмент и большой С-концевой фрагмент, который называется кленовским фрагментом. Последний сохраняет 3’-нуклеазную и полимеразную активность и часто используется для синтеза ДНК in vitro.
(3’®5’)-экзонуклеазный домен, ответственный за корректорскую активность ДНК-полимеразы I, не является обязательным и отсутствует у бактерий из родов Thermus и Rickettsia. Остальные два домена жизненно важны для клеток и обеспечивают участие ДНК-полимеразы I в процессах репарации и удаления рибонуклеотидной части фрагментов Оказаки в отстающей нити. Поэтому делеция гена polA летальна для E. coli, а нелетальные мутации понижают в 10 раз скорость соединения фрагментов Оказаки и повыщаютповещают чувствительность клеток к агентам, повреждающим ДНК.
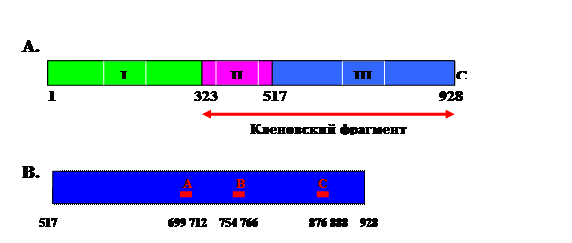
Рис. 1.6. Доменная с
труктура ДНК-полимеразы I E.coli.
A – взаимное расположение доменов. I – 5’-нуклеазный домен, II – (3’®5’)-экзонуклеазный домен, III – ДНК-полимеразный домен. Стрелка соответствует кленовскому фрагменту ДНК-полимеразы I.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
B – мотивы полимеразного домена, консервативные среди бактериальных ДНК-полимераз I.
Эти мотивы имеют следующие консенсусные последовательности: 699hhhh D hhxEhx712 (A), 754 R pxx K xxxhGhh Y 766(B) и 876hhxhH D ЕhxxЕ888 (С) -, где h – гидрофобный остаток, х – произвольный остаток, р – преимущественно R (нумерация остатков ДНК-полимеразы I E. coli)
Полимеразные домены различных ДНК-полимераз I и ДНК-полимеразы фага Т7 очень похожи друг на друга и содержат 6 консервативных участков, среди которых три аналогичны консервативным мотивам А, В и С главного семейства В эукариотических ДНК-полимераз и играют наиболее важную роль во время синтеза ДНК. Мотивы А и С ДНК-полимеразы I E. coli содержат кислые остатки асп705 и асп882 соответственно, связывающие катионы Mg+ в каталитическом активном центре (см. рис. 1.1В), а мотив В включает инвариантный положительно заряженный остаток (арг758) и ароматический остаток (тир766). Эти три мотива являются элементами полости, с которой связывается включающийся дНТФ в активном полимеразном центре.
- 
Рис. 1.7. Модель трехмерной структуры полимеразного домена ДНК-полимеразы I E.coli.
Указаны положения, в которых начинаются консервативные мотивы А, В и С
Рентгеноструктурный анализ кленовского фрагмента ДНК-полимеразы I E. coli впервые обнаружил специфическую архитектуру полимеразного домена, который по форме напоминает кисть правой руки (рис. 1.7) и состоит из пространственных доменов «ладони» (palm), «большого пальца» (thumb) и «пальцев» (fingers). Аналогичную трехмерную организацию имеют и все изученные экспериментально полимеразные домены других ДНК-полимераз и обратной транскриптазы вируса иммунодефицита человека. Можно предположить, что так же организованы и полимеразные домены большинства ДНК-полимераз. Наиболее консервативна топология домена ладони, состоящего из b-слоя, фланкированного двумя a-спиралями. В домене ладони расположены консервативные мотивы А и С. Домены большого пальца и пальцев у ДНК-полимеразы I E. coli образованы a-спиралями, причем спираль О домена пальцев совпадает с мотивом В. У других ДНК-полимераз организация этих доменов гораздо менее консервативна. Хотя домен больших пальцев остается преимущественно a-спиральным, но тонкие детали его топологии неодинаковы. Еще более велики различия доменов пальцев. В целом, для архитектуры полимеразного домена характерно существование внутренней полости для связывания матрицы-затравки ДНК и входящего дНТФ, основанием которой является домен ладони.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
На примере бактериальной ДНК-полимеразы Iмы рассмотримпоследовательные изменения конформации этой полости во время полимеразного каталитического цикла. Эти рентгеноструктурные данные были получены для нескольких свободных бактериальных полимераз и их комплексов с ДНК в присутствии и в отсутствие дНТФ. Вероятно, такие же изменения происходят во время работы других ДНК-полимераз. «Рабочий цикл» состоит из 4 последовательных стадий: 1) связывания ДНК-полимеразы с матрицей-затравкой ДНК; 2) связывания с комплексом полимераза-ДНК подходящего дНТФ, комплементарного первому остатку матрицы; 3) нуклеофильной атаки, приводящей к образованию фосфодиэфирной связи; 4) освобождения пирофосфата. Первые две стадии протекают очень быстро, и общую скорость полимеризации лимитирует стадия 3 или предшествующее ей изменение конформации.
Первая стадия сопровождается конформационными изменениями домена большого пальца, который поворачивается в сторону домена ладони. Одновременно изменяется положение спиралей Н1 и Н2 на конце большого пальца, которые сближаются с днДНК. В результате из домена большого пальца и прилежащей части домена ладони образуется цилиндрический канал диаметром 30 Å, охватывающий дуплекс ДНК. В нем аминокислотные остатки ДНК-полимеразы контактируют с ДНК по малой канавке. Взаимодействия с ДНК полимеразой вызывают изменения конформации как в днДНК (изгиб на расстоянии 3 п.н. от праймерного конца), так и однонитевой затравке, первый нуклеотид которой поворачивается на угол > 90о, а следующие основания – на 180о. Этот переход матричной нити в S-образную конфигурацию смещает первый матричный нуклеотид с оси дуплекса ДНК, на которой вместо него оказывается остаток тир766 спирали О домена пальцев.
Второе изменение конформации происходит после связывания дНТФ и в основном затрагивает домен пальцев и смежную область домена ладони. Домен пальцев поворачивается в сторону 3’-конца затравки, а его О-спираль с мотивом В вращается на 40о в сторону двух остатков асп каталитического центра. В результате сайт связывания дНТФ переходит из «открытой» формы в «закрытую». Первый остаток матрицы возвращается на ось днДНК, вытесняя с нее остаток Y766, сохраняющий стэкинг-взаимодействие с этим матричным нуклеотидом. В «закрытой» форме входящий дНТФ образует уотсон-криковскую пару с первым матричным основанием. В стабилизации этой пары участвуют также взаимодействие ароматических колец остатков фен и тир с мотиве В с азотистым основанием дНТФ и гидрофобные взаимодействия боковых цепей остатков или и глу с дезоксирибозой. Положительно заряженный остаток арг758 мотива В взаимодействует с отрицательно заряженным трифосфатом дНТФ, а остатки асп705 и асп882 активного центра в домене ладони при участии двух катионов Mg2+ устанавливают правильную геометрию трифосфата дНТФ, необходимую для нуклеофильной атаки 3’-концом затравки (см. рис. 1.1В). Таким образом, замкнутая структура белка, удерживающая в правильной конфигурации пару дНТФ-матричное основание, создается конформационными изменениями ДНК полимеразы в соответствии с моделью ииндуцированногоиндуцированного соответствия.
После включения очередного нуклеотида в растущую цепь ДНК полимераза возвращается в «открытую» форму, транслоцируется к следующему положению матрицы и освобождает пирофосфат. В освобождении пирофосфата важную роль играет взаимодействие остатков арг и лиз в О-спирали с a- и b-фосфатными группами. Изменение положения этих основных остатков во время перехода в открытую форму способствует удалению пирофосфата из каталитического центра и предотвращает обратную реакцию пирофосфоролиза ДНК.
|
|