Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
III Водорастворимые 4 страница
|
|
hormaino—побуждаю, привожу в движение) было предложено Бейлиссом и Стерлингом (1905). В настоящее время гормонами называют вещества, образующиеся в железистых клетках, выделяющиеся в кровь или лимфу и регулирующие обмен веществ и развитие организма.
Гормонам присущи следующие общие биологические признаки:
1) дистантность действия, т. е. они регулируют обмен и функции эф- фекторкых клеток на расстоянии,
2) строгая специфичность биологического действия, т. е. один гормон нельзя целиком заменить другим;
3) высокая биологическая активность — достаточно очень малых количеств^, порой десятка мнкрограмм, чтобы сохранить жизнь организма.
мически связанные с отделами центральной нервной системы, и периферические (табл. 33).
Кроме перечисленных желез эндокринной функцией обладают и другие клетки^ которые выделяют биологически активные вещества, похожие по своим свойствам на гормоны. Поэтому их принято называть еормоноподоб- ными веществами или гормоноидами (местные гормоны, парагормоны). Они, как правило, действуют в месте своего образования и выделяются клетками,
Таблица 33 Гормоны эндокринных желез
|
Продолжение табл 33
|
рассеянными в разных органах. Выделяются гормоноиды клетками желудочно-кишечного тракта (они регулируют процессы пищеварения), энтерохрома- финными клетками кишечника (выделяют серотонин, регулирующий функцию кишечника), тучными клетками соединительной ткани (гепарин, гистамин).
клетками почек, семенных пузырьков и других органов (простагландины) и т. д. Возможно, эти эндокринные клетки являются своеобразным представительством эндокринной системы на периферии. Это подтверждается обнаружением некоторых истинных гормонов в периферических органах; например соматостатина в поджелудочной железе, в печени и т. д.
По химическому строению гоомо- ны подразделяются на:
1) белково-пептидные (гормоны гипоталамуса, гипофиза, поджелудочной и паращитовидной желез, каль- циотокнн щитовидной железы);
2) производные аминокислот (адреналин — производное аминокислот фенилаланина и тирозина; иодтиронины — производные тирозина; мелато- нин — производное трнптофана);
3) стероиды (половые гормоны — андрогены, эстрогены и гестагены; кортикостероиды)
2. Схема нейроэндокринных взаимосвязей
Поток информации о состоянии внешней и внутренней среды организма поступает в нервную систему, где перерабатывается, а в ответ посылаются регуляторные сигналы к периферическим тканям и органам. Причем нервная' регуляция обмена веществ и функций исполнительных органов осуществляется не только благодаря поступлению нервных импульсов по центростремительным нервам, но и опосредованно, через эндокринную систему. Объединяются оба потока информации — нервной и гормональной — на уровне гипоталамуса. Нервные импульсы, поступающие от различных отделов головного мозга, влияют на секрецию клетками гипоталамуса нейропептидов, которые регулируют выделение тропных гормонов гипофиза, а последние влияют на секрецию гормонов в периферических железах.
Образование и секреция гормонов периферическими железами происходят непрерывно. Это необходимо для поддержания необходимого урорня их в крови, поскольку они быстро инактивируются и выводятся из организма Требуемый уровень гормона в крови поддерживается благодаря механизму- саморегуляции. В основе его лежат межгормональные взаимоотношения, которые носят название «плюс — минус» взаимодействий или взаимопротиворечивых отношений

|
Схема этих взаимоотношений показана на рис. 74. Тропные гормоны стимулируют образование и секрецию гормонов периферическими железами
(знак «плюс»), а последние по механизму отрицательной обратной связи угнетают (знак «минус») образование тропных гормонов, действуя через клетки гипофиза (короткая обратная связь, рис. 74, 1) или нейросекретор- ные клетки гипоталамуса (длинная обратная связь, рис. 74, -2). В последнем случае угнетается секреция либеринов в гипоталамусе. Кроме того, существует метаболитно-гормональная обратная связь (рис. 74, 5): гррмон, действуя на обмен веществ в тканях, вызывает изменение содержания какого-либо метаболита в крови, а тот, по механизму обратной связи, влияет на секрецию гормонов в периферических железах (или непосредственно, или через гипофиз и гипоталамус). На роль метаболитов — индикаторов действия гормонов на клетку могут претендовать глюкоза (индикатор состояния углеводного обмена), аминокислоты (индикатор состояния белкового обмена), нуклейчи- ды и нуклеозиды (индикатор состояния нуклеинового и, видимо, белкового обмена), жирные кислоты и холестерин (индикатор состояния липидного обмена), Са2*, Na+, К+, С1_ (возможно, и другие ионы) и вода (индикатор состояния водно-солевого баланса). Для некоторых нз упомянутых метаболитов (глюкоза, некоторые аминокислоты, Са2+, Na+, К+, С1—, Н20) уже доказано, что они могут по механизму обратной связи регулировать секрецию гормонов.
3. Общие представления о действии гормонов
Гормоны, секретируемые железами, обычно связываются в крови со специфическими транспортными белками плазмы (или в некоторых случаях адсорбируются на клетках крови) и доставляются к периферическим тканям, где оказывают влияние на их обмен веществ и функцию. Не все ткани одинаково реагируют на действие гормона. Тканн (клетки, органы) с высокой чувствительностью к гормонам, т. е. ткани, в которых гормоны вызывают наиболее выраженные сдвиги в обмене и функции, называют «мишенями» для данного гормона, а остальные относят к тканям-«немишеням», хотя это разделение условно. Влияние гормонов простирается буквально на все стороны обмена веществ. Однако, чтобы понять механизм действия любого гормона, как, впрочем, и любого внеклеточного регулятора, необходимо рассмотреть общие принципы гормональной регуляции обмена веществ.
Различают следующие возможные варианты действия внеклеточных регуляторов, s том числе и гормонов:
1) мембранный, или локальный;
2) мембранно-внутриклеточный, или косвенный;
3) цитозольный, или прямой.
Мембранный тип действия гормонов
Мембранный тип действия заключается в том, что гормон в месте связывания с клеточной мембраной изменяет проницаемость для глюкозы, аминокислот, некоторых ионов. В этом случае гормон выступает как аллостерическнй эффектор чранспортных систем мембран. Поступление глюкозы и аминокислот оказывает, в свою очередь, влияние на биохимические процессы в клетке, а изменение распределения ионов по обе стороны мембраны влияет на электрический потенциал и функцию клеток. Мембранный тип действия гормонов редко встречается в изолированном виде. В качестве примера можно назвать инсулин, который обладает как мембранным (вызывает местные изменения транспорта некоторых ионов, глюкозы, возможно, аминокислот), так и мембранно-внутриклеточным типом действия.
Мембранно-внутриклеточный механизм регуляции обмена веществ
Мембранно-внутриклеточный тип действия характерен для гормонов (и других внеклеточных регуляторов), которые не проникают в клетку и поэтому влияют на обмен веществ через внутриклеточный химический посредник (вторичный посредник), который и является своеобразным полномочным представителем гормона внутри клетки.
Оказалось, что открытый в 1957 г. Сазерлендом 3', 5'-АМФ, или цАМФ, является внутриклеточным посредником гормонов и других внеклеточных регуляторов. В настоящее время на роль внутриклеточных посредников претендуют три группы веществ: циклические нуклеотиды — цАМФ и цГМФ, ионы Са2+ и 2', 5'-олиго (А)„-адениловый олигонуклеотид. Гормоны (или другие внеклеточные регуляторы) регулируют образование вторичных посредников в клетке, а те, в свою очередь, влияют на активность и количество разных ферментов и тем самым изменяют биохимические функции клетки.
Для гормона (или любого другого внеклеточного регулятора) мембранные рецепторы — своего рода «кнопки> на пульте управления клеточным метаболизмом. Через них внеклеточный регулятор влияет на функцию сигнальных систем (обычно это ферменты), запускающих образование или поступление внутриклеточных посредников.
Регуляция обмена через циклические нуклеотиды. Внеклеточные регуляторы действуют на образование циклических нуклеотидов через одну из двух сигнальных систем: адёнилатциклазу или гуанилатциклазу. Первая контролирует образование цАМФ, вторая — цГМФ:
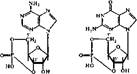
|
| циклический J'.5" -AM4> |
| циклический 3'.5'-ГМФ |
Аденилатциклаза встроена в мембрану и состоит из трех взаимосвязанных частей (рис. 75):
1) узнаюшей (R), представленной набором мембранных рецепторов и находящейся снаружи;
2) сопрягающей (N), представленной особым белком (называется N-бел- ком), который занимает промежуточное положение в липидном слое мембраны
 Рис. 75. Схема мембран но-внутриклеточного механизма действия гормонов на обмен веществ клеток (посредством циклических нуклеотидов)
Рис. 75. Схема мембран но-внутриклеточного механизма действия гормонов на обмен веществ клеток (посредством циклических нуклеотидов)
|
между рецептором и катаболической частью. N-Белок имеет участок связывания и расщепления ГТФ;
3) каталитической (С), являющейся ферментным белком, т. е. собственно аденилатциклазой (активный центр ее обращен внутрь клеткн, благодаря чему находящийся здесь АТФ может превращаться в цАМФ по схеме: АТФ-> 3', 5'-АМФ + Н< Р207).
Как же работает аденнлагцнклазная система? Как только гормон связывается с рецептором, то комплекс гормон — рецептор взаимодействует с N-белком. В результате изменяется конфигурация N-белка и происходит связывание ГТФ с этим белком, а правильнее сказать, замещение ГДФ, находящегося в неактивном N-белке, на ГТФ. Комплекс N-белок — ГТФ является аллостерическим активатором собственно аденилатииклазы (каталитической части). Активация аденилатциклазы приводит к наработке цАМФ внутри клетки из АТФ.
В активном состоянии аденилатииклаза поддерживается до тех пор, пока существует комплекс гормон --рецептор, который обеспечивает образование комплекса N-белок — ГТФ, активирующего аденилатциклазу. При диссоциаций комплекса гормон — рецептор прекращается его действие на N-белок, ГТФ расщепляется до ГДФ и Фн, а остающийся комплекс N-белок — ГДФ не способен активировать аденилатциклазу. В результате прекращается наработка цАМФ.
Благодаря такому устройству аденилатциклазной системы происходит многократное усиление гормонального сигнала. Даже единственная молекула связавшегося с рецептором гормона заставляет работать аденилатциклазу (пусть в течение короткого времени), которая, как всякий фермент, обеспечивает быструю наработку цАМФ. Происходит как минимум 10—100-кратное усиление сигнала; на одну молекулу связавшегося гормона образуется 10—100 молекул цАМФ внутри клетки.
Сходным образом гормоны и другие внеклеточные регулятиры запускают образование другого внутриклеточного посредника — цГМФ. В этом случае комплекс гормон — мембранный рецептор склонен активировать гуа- нилатциклазу (см. рис: 75). В отличие от аденилатциклазы она непрочно связана с клеточной мембраной, поэтому контактирующий с ней рецептор глубже погружается в липидный слой мембраны, чтобы контактировать с гуанилатциклазой. При активировании гуанилатциклазы нарабатывается цГМФ из ГТФ.
В клетке обнаружена и растворимая в цитоплазме гуанилатциклаза, которая, очевидно, реагирует на изменения внутриклеточной среды. (Возможно, имеются циклазы для образования и других циклических нуклеотидов типа цДМФ и цУТФ, но это пока точно не доказано.)
Циклические нуклеотиды далее активируют протеинкнназы, локализованные во всех отсеках цитоплазмы и ядре. Существует не менее двух групп протеинкиназ: цАМФ-зависимые и цГМФ-зависимые, которые активируются своим циклическим нуклеотидом. цАМФ-Протеинкиназы состоят из четырех субъединиц: из двух регуляторных (R), с которыми связываются циклические нуклеотиды, и двух каталитических (С). Присоединение к R-субъ- единнцам цАМФ способствует диссоциации тетрамера; при этом С-субъеди-- ницы объединяются в димер, который и является активной формой протеинкиназы.
Предполагается, что ц ГМФ-протеинкиназа является димером. На каждой субъединице имеется каталитический центр (С) и участок связывания цГМФ (R). Для активирования протеинкиназы необходимо связывание двух молекул цГМФ с R-участками,
Роль активированных протеинкиназ сводится к фосфорилированию белков по схеме
Белок + АТФ Прог< |, н*" " азы Фосфопротеид + АДФ
Разные протеинкиназы фосфорилируют разные белки. Поэтому, казалось бы, однообразная протеинкиназная реакция вызывает целую гамму разнообразных биологических эффектов, поскольку функции, выполняемые конкретными белками, разные. Они, по сути дела, определяют жизнедеятельность клетки в целом. Функция одних белков после фосфорилирования их протеинки- назами активируется, функция других угнетается.
К тому же одна группа функциональных белков подчиняется регулирующему влиянию цАМФ, а другая—цГМФ. Поэтому в зависимости от мембранного рецептора, связывающего гормон, включаются или цАМФ-за- висимые биологические процессы, или цГМФ-завиеимые процессы (см. рис. 75). Причем часто цАМФ и цГМФ действуют противоположно, поэтому представляется уникальная возможность для проявления противоположного действия одним и тем же внеклеточным регулятором на обмен веществ в разных клетках, если в них имеются разные типы рецепторов.
Например, адреналин связывается как с а-, так и с p-адренорецепторами, но p-адренорецепторы включают аденилатциклазу и образование цАМФ, а а-адренорецепторы — гуанилатциклазу и образование цГМФ. Следовательно, биохимические функции в клетках, содержащих разные адренорецепторы, будут изменяться по-разному, несмотря на то, что на них влияет одни и тот же внеклеточный регулятор.
Каким же образом прекращается влияние циклических нуклеотидов кз биохимические процессы? Этой цели служат специальные ферменты, разрушающие и сами циклические нуклеотиды, и результат • их действия — образовавшиеся фосфо протеиды. Гидролиз циклических нуклеотидов осуществляется фосфодиэстеразами по схеме
3', 5'-АМФ (или 3', 5'-ГМФ) -ТЩ? ±М__-»5'.АМФ (или б'-ГМФ)
Образующиеся просто АМФ и ГМФ не способны активировать протеин- киназы. Кроме фосфодиэстераз в клетках имеется фосфопротеидфосфатаза, которая гидролизует фосфопротеид по схеме
Фосфопротенд + HjO -»- Белок + Н3РО4
Оба фермента — фосфодиэстераз а и фосфопротеидфосфатаза — полностью снимают влияние на обмен веществ, вызванное действием внеклеточных регуляторов.
2', 5'-Олиго(А) как внутриклеточный посредник. Роль этого аденилового олигонуклеотида как внутриклеточного регулятора обмена'веществ мало изучена. Образование его осуществляется специальным ферментом — олиго(А)- синтетазой, активность которой может меняться под действием внеклеточных регуляторов.
Механизм регуляции обмена посредством ионов Са2+. Необычная роль ионов Са2+ как внутриклеточных посредников внеклеточных регуляторов выявилась относительно недавно. В отличие от других посредников ионы Са2+ не могут превращаться, поэтому кальциевые сигнальные системы мембран, реагирующие на внешние стимулы, способны лишь изменять поступление Са8+ в цитоплазму. Внутриклеточное содержание Са2+ ничтожно — К)" 7, моль/л, тогда как вне клетки 10~3. Ионы Са2+ поступают из внешней. среды по двум «кальциевым каналам» в мембране. Поток Са2+ регулируется Са2+-АТФазои клеточной мембраны, которая за счет энергии АТФ откачивает Са2+ в обмен на Na+ из цитоплазмы во внешнюю среду. Внутри клетки ионы Са2+ депонируются в матриксе митохондрий, а в мышечной ткани — в цистернах (замкнутых пузырьках) саркоплазматнческого ретикулума.
Кальций, поступающий из внешней среды или внутриклеточных-депо под действием различных стимулов, взаимодействует с Са-связывающими белками цитоплазмы, выполняющими роль регуляторов. Таким Са-связывающим белком является калмодулин (КМ). Комплекс Са2+ • КМ изменяет (модулирует) активность разных ферментов, что ведет к изменению биохимических функций клеток. В целом регуляция посредством Са" + происходит примерно по.следующей схеме:
внеклеточный фактор — мембранные рецепторы — сигнальные кальциевые системы (Са2+-АТФазы) — поступление Са2+ в цитоплазму — образование регуляторного комплекса Саг+ • КМ — изменение активности ферментов — изменение клеточных функций
^ Таким образом, чувствительность тканей и органов к внеклеточным ре-
специфическое регуляторное влияние определяется тем внутриклеточным посредником, через- который гормон-рецепторный комплекс преимущественно влияет на обмен веществ (табл. 34).
Кроме природных веществ, действующих через аденилат- и гуанилэтцик- лазы на образование цАМФ и цГМФ, объектом возможного воздействия лекарств является фосфодиэстераза. Если препараты проникают внутрь клетки и ингибируют одну из фосфодиэстераз цАМФ или цГМФ, то они способны имитировать эффект, оказываемый на обмен веществ природным гормоном и медиаторами, действующими через циклические нуклеотиды. Активаторы фосфодиэстеразы снижают действие этих же гормонов и медиаторов. В медицине пользуются этой возможностью, применяя препараты — ингибиторы фосфодиэстеразы. К ним относятся производные ксантинов: кофеин, теофиллин, теобромин, эуфиллин и др. Эти препараты повышают содержание преимущественно цАМФ, а такой препарат, как трентал, — преимущественно цГМФ.
Цитозольный механизм действия
Цитозольный тип действия характерен для гормонов, способных проникать через липидный слой плазматической мембраны, т. е. по своим физико- химическим свойствам относящихся к липофнльным веществам, например для стероидных гормонов (витамин D тоже близок по. типу действия'на обмен веществ к стероидным гормонам, поэтому его часто относят к гормональным веществам). Гормоны с цитозольным типом действия проникают внутрь клетки, где вступают в комплекс с цнтозольными рецепторами. В комплексе с рецептором гормоны регулируют количество ферментов в клетке, избирательно влияя на активность генов хромосом ядра, и тем самым изменяют обмен веществ и функции клетки. Поскольку проникший в клетку гормон сам участвует в механизмах регуляции количества ферментов, то подобный тип действия называется также прямым в отличие от мембран- но-внутриклеточного, когда гормон регулирует обмен веществ лишь косвенно, через внутриклеточные посредники. Иодтиронины по лнпофиль- ности занимают промежуточное положение между стероидами и остальными водорастворимыми гормонами. Возможно поэтому они обладают смешанным типом действия на обмен веществ клеток, т. е. мембранно-внутрнкле- точным и цитозольным.
Схематически цитозольный механизм показан на рис. 76. Стероидные гормоны и иодтиронины, проникая через клеточную мембрану, связы- 380
регуляторов, действующих через внутриклеточных
посредников
Вешествэ, действующие через яАМФ
Гормоны гипофиза: тиреотролин. кор- тнкотропин, соматотропин, гонадотро- пнны, p-липотропин, меланотропнн, вазопрессин
Гормоны эпифиза (мелатоннн) Гормоны щитовидной железы: тироксин, трииадтироннн Гормоны паращнтовндных желез: па- ратнрин
Гормоны поджелудочной железы: глюка гон
| Кальииотонин Ка дышат он ик (?) Инсулин |
Гормоны мозгового вещества надпочечников: адреналин (через р-адрено- рецепторы)
Адреналин (через fl-адренорецепторы) Дофамин
Гистамнн (через Н2-реиепторы)
Н ей ромеди агоры
Адреналин (через а-ад- I ренорецепторы) Норадреналин (через о - адреноре цептор ы) Ацетилхолин. гистамин (через Н|-рецепторы).
Тканевые регуляторы деления кл
Фитогемаглютнинны
| Кейлокы Интерферон |
Фактор роста нервов
| Гор моноиды I Простагландин Fa, |
| Простагландины Б,, Ег Гастрин |
Холерный токсин
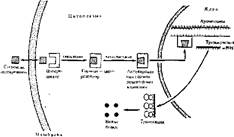 Рис 76. Схема цнтозольного механизма действия гормонов
Рис 76. Схема цнтозольного механизма действия гормонов
|
ваются с цитозольными рецепторами, через которые гормоны оказывают регуляторное действие на метаболизм клеток. Циторецепторы являются белками с молекулярной массой 60 000—120 000. Для них характерна стерео- специфичность связывания, а отсюда высокое сродство к своему гормону.
Первичный комплекс гормон — циторецептор подвергается в цитоплазме активации, которая заключается в перестройке молекулы цитореиептора. В «активированной» форме гормон-рецепторный комплекс способен проникать через ядерную мембрану к хромосомам -ядра и взаимодействовать с ними! Конкретный гормон-рецепторный комплекс, связываясь с регуляторнымк белками хроматина (гистоны, негистоновые белки) или с ДНК, регулирует или деление клеток, или транскрипцию «своих» генов в неделящихся клетках и синтез специфических белков. Этим определяется специфический эффект каждого гормона.
Поскольку стероидные гормоны влияют только на активность генов в хромосомах, то для них более характерна, чем для гормонов с мембранно- внутриклеточным типом действия, регуляция роста и Дифференцировки клеток, т. е. влияние на развитие организма. При введении таких гормонов в больших дозах может проявиться и прямое их действие на ферменты и внутриклеточные мембраны, так как для связывания массивных количеств проникшего гормона не хватает цитозольных рецепторов.
4. Получение и практическое применение гормонов
Для практического применения гормоны получают извлечением из биологического материала, химическим синтезом и методами генной инженерии. Первым способом получают инсулин и глюкагон из поджелудочных желез, корти- 382 котропин и меланотропии из гипофизов крупного рогатого скота, фоллитропин (сывороточный гонадотропин) из сыворотки крови и лютропин (хориониче- ский гонадотропин) из мочи жеребых кобыл. Путем химического синтеза в настоящее время получают все стероидные гормоны, их аналоги и производные, иодтиронины и другие гормоны — производные аминокислот, пептидные гормоны типа окситоцина. Лабораторным синтезом получены практически все белковые гормоны. Методом генной инженерии получены в лабораторных условиях инсулин, соматостатин и др.
В медицинской практике гормоны применяются для заместительного и патогенетического лечения. В последнем случае используются отдельные свойства гормонов (противовоспалительные, анаболические и др.), хотя их содержание в организме больного человека не снижено.
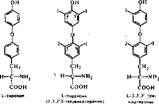
5. Гормоны щитовидной. железы
Щитовидная железа секретирует две -группы гормонов с разным влиянием на обмен веществ. Иодтиронины — тироксин и трииодтиронин регулируют
|
энергетический обмен и влияют на деление и дифференцировку клеток, определяя развитие организма. Кальциотонин (белок с молекулярной массой около 30*000) регулирует фосфорно-кальциевый обмен; его действие удобнее рассматривать вместе с гормонами паращитовидных желез.
|
|