Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Для новых пользователей первый месяц бесплатно. Чат-бот для мастеров и специалистов, который упрощает ведение записей: — Сам записывает клиентов и напоминает им о визите;
— Персонализирует скидки, чаевые, кэшбэк и предоплаты;
— Увеличивает доходимость и помогает больше зарабатывать; Начать пользоваться сервисом

Плазменные белки гемостаза
|
|
Плазменные белки гемостаза образуют 2 ферментативные системы крови, имеющие своей целью поддержание гемостатического баланса (рис. 31):
1. Система свертывания плазмы. Система со
стоит из ферментов, неферментативных бел
ковых катализаторов (кофакторов) и инги
биторов свертывания. Конечной целью этой
системы является образование важнейшего
фермента тромбина, а в конечном итоге -
фибринового сгустка, составляющего осно
ву гемостатического тромба.
2. Система фибринолиза. Конечной целью этой
системы является образование главного фер
мента фибринолиза плазмина и лизис фибри
нового сгустка. Эту систему составляют плаз-
миноген и его активаторы и ингибиторы.
Обе эти системы имеют сходные черты:
• В обеих системах происходит многоэтапный ферментативный процесс актива-
ции, в котором участвует ряд белков - про-теаз.
• По крайней мере, in vitro имеется несколько
путей запуска каждого процесса, а в итоге об
разуется один конечный продукт.
• Многие реакции нуждаются в наличии спе
цифической поверхности и ионов кальция.
In vivo твердой фазой для фиксации реаги
рующих белков служат кислые фосфолипи-
ды клеточных мембран, в частности мемб
ран тромбоцитов, фибробластов, возможно
лейкоцитов.
Кроме каскадных систем свертывания плазмы и фибринолиза, к плазменным белкам гемостаза относятся многочисленные ингибиторы и активаторы, эффекты которых проявляются как действие антикоагулянтов или прокоагулянтов и соответственно ингибиторов или активаторов фибринолиза.
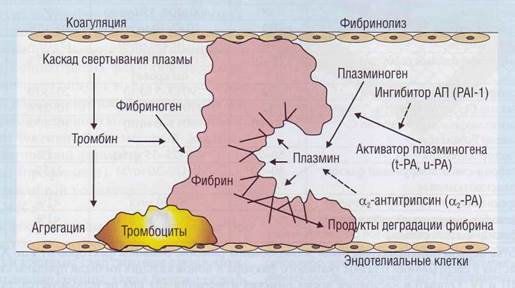
Рис. 31. Система свертывания крови и система фибринолиза - каскадные протеолитические ферментативные системы, обеспечивающие гемостатический баланс крови
Плазменные белки гемостаза
Система свертывания плазмы
Система свертывания плазмы - ферментативная система, осуществляющая каскад протео-литических реакций, в результате которых происходит образование фибриновой пробки в месте повреждения сосуда. Система свертывания тесно связана с другими протеолитическими системами плазмы, в том числе с системой фибри-нолиза. Белки свертывания плазмы, входящие в каскад свертывания крови, принято называть термином «фактор». В соответствии с международной номенклатурой факторы свертывания плазмы обозначаются римскими цифрами (табл. 6). Активные формы факторов обозначаются теми же римскими цифрами, но с добавлением аббревиатуры «а».
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Практически все факторы системы свертывания крови циркулируют в кровотоке в форме неактивных проэнзимов или в форме неактивных кофакторов. Исключение составляет фак-
тор VII, примерно 1-2% которого в норме циркулируют в активной форме. При запуске свертывания крови происходит каскадная активация проэнзимов и кофакторов (рис. 32). Процесс активации представляет собой ограниченный протеолиз неактивных предшественников до активных энзимов и кофакторов. Активированные энзимы являются сериновыми протеа-зами (за исключением фактора XIII). Активированные кофакторы, не обладая самостоятельной ферментативной активностью, играют роль коферментов.
Сериновыми протеазами являются активированные факторы II, VII, IX, X, XI, XII, ПК.
Трансглютаминаза - фактор XIII.
Кофакторы - факторы V, VIII, ВМК.
Содержание компонентов гемостаза, в том числе плазменных факторов свертывания, в системе циркуляции существенно больше, чем необходимо
| Таблица 6 |
| Плазменные факторы свертывания крови |
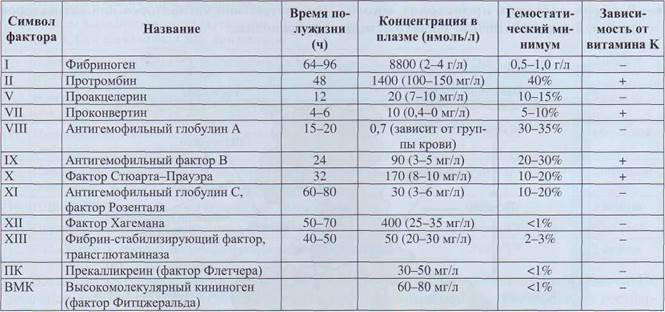
При разработке первой номенклатуры были использованы римские символы факторов от I до XIII. Для обозначения участия в свертывании плазмы тканевого фактора и ионов кальция им были приданы символы соответственно III и IV. Однако в настоящее время римская нумерация для них не используется, так как они не относятся к плазменным факторам свертывания (тканевой фактор - это тканевой компонент вне сосудистой системы, ионы Са не являются белком). Фактор VI в классификации не употребляется, так этим символом ошибочно был назван фактор Va.
Плазменные белки гемостаза
|
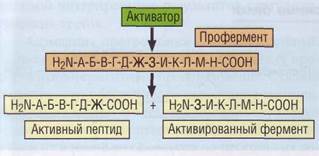
Рис. 32. Протеолитическая активация факторов гемостаза. Путем ограниченного протеолиза из неактивного предшественника образуются активный пептид и активированный фермент
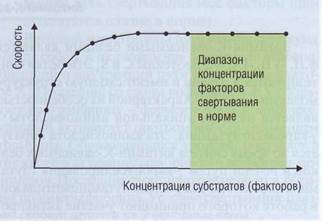
для формирования фибринового сгустка. Процесс свертывания происходит в условиях насыщения субстратами (рис. 33). Вследствие этого образование гемостатического тромба может быть достигнуто при значительном диапазоне концентрации и активности конкретного фактора свертывания. Клинические проявления недостаточности компонентов свертывания возникают при их существенном уменьшении, если обратиться к рис. 33 - то это начальный диапазон, при котором скорость реакции зависит от концентрации фактора.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
Для эффективного взаимодействия и активации белков свертывания крови необходимо образование комплексов этих белков, их кофакторов и субстрата (рис. 34). Эти условия не могут возникнуть в жидкой фазе. Поэтому большинство процессов активации промежуточных факторов свертывания протекают на фосфолипидах клеточных мембран. В месте сборки комплексов происходит концентрация факторов свертывания. Здесь же присутствуют кофакторы, которые существенно ускоряют процесс формирования сгустка. В создании активного комплекса участвуют:
• Фермент (активный плазменный фактор -
протеолитический фермент).
• Субстрат (профермент).
• Активированный кофактор.
• Ионы Са (Са2+).
• Кислые фосфолипиды и специфические ре
цепторы на поверхности клеток.
Все белки системы свертывания крови можно разделить на две группы. Одни белки для полноценного формирования требуют наличия витамина К (витамин-К-зависимые белки), а другие - нет.
Рис. 33. Соотношение между концентрацией факторов и скоростью процесса свертывания. В норме скорость коагуляции практически не определяется концентрацией факторов, так как они присутствуют в избытке и процесс идет в состоянии насыщения. Только после значительного истощения фактора его концентрация будет влиять на скорость реакции и соответственно на скорость свертывания плазмы
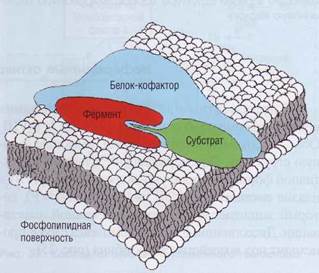
Рис. 34. Модель сборки комплекса факторов свертывания крови. На поверхность твердой фазы (фосфолипиды фибробластов, макрофагов, активированных тромбоцитов либо, в патологических ситуациях, мембраны поврежденных клеток, бактерий и др,) прикрепляется (интернали-зуется) крупный кофакторный белок, который организует место контакта факторов свертывания, те в свою очередь взаимодействуют друг с другом по принципу комплементарности
Плазменные белки гемостаза
Витамин-К-зависимые белки
|
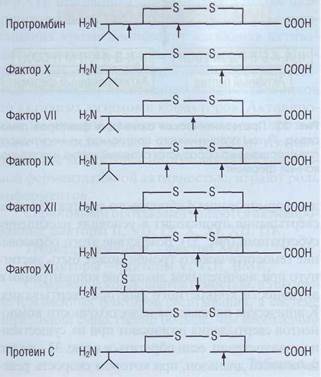
Витамин-К-зависимыми белками являются ф.II, -VII, -IX, -X, протеины С и S. Эти белки синтезируются в печени и имеют сходную структуру молекулы (рис. 35). Характерной их особенностью является наличие уникальной аминокислоты -у-карбоксиглутамина. Эта аминокислота образуется во время синтеза витамин-К-зависимых белков в печени путем у-карбоксилирования глута-мина ферментом у-карбоксиглутаминпептидазой, в работе которого принимают участие активированные формы витамина К (рис. 142). у-карбок-сиглутамин дает возможность витамин-К-зависи-мым белкам с помощью ионов Са2+ образовывать комплексы с кислыми фосфолипидами.
Рис. 35. Структурная организация некоторых плазменных белков системы гемостаза. Стрелками показаны места протеолитического гидролиза, в результате которого происходит переход неактивных проферментов в активные ферменты - сериновые протеазы каскада коагуляции. Двузубцем обозначены витамин-К-зависимые факторы, имеющие в своей структуре карбоксилированную глюта-миновую кислоту
Неферментные активаторы свертывания крови
К неферментным активаторам свертывания крови (коферментам) относятся факторы V и VIII. Оба - высокомолекулярные белки, имеющие сходную структуру. Они циркулируют в плазме в неактивной форме и активируются тромбином. ф.VIII в плазме связан с фактором Виллебранда (vWF), который защищает его от преждевременной инактивации. Диссоциация фVIII из комплекса с vWF происходит под воздействием тромбина (рис. 25).
ф.Va и -Villa образуют на фосфолипидных мембранах комплексы с ф.Ха и -IХа соответственно. Специфическая активность ф.Ха и -IХа в комплексах с кофакторами в десятки тысяч раз больше, чем изолированных. Основным ингибитором ф.Va и -VIIIa является комплекс протеин С - протеин S.
Классический коагуляционный каскад активации тромбина
Изучение процесса свертывания крови до настоящего времени происходит в основном in vitro в смоделированных условиях. Исследование взаимодействия плазменных белков гемостаза в отрыве от других компонентов привело к созданию так называемой «классической» теории коагуля-ционного каскада активации тромбина. В насто-
ящее время эта теория пересмотрена с учетом вновь полученной информации о взаимодействии различных компонентов гемостаза. Однако базовые принципы изложенной ниже классической каскадной теории считаются верными до настоящего времени. Кроме того, знание классического каскада свертывания крови необходимо для пра-
Плазменные белки гемостаза
вильнои интерпретации результатов коагулоло-гических тестов.
Активация протромбина - многостадийный процесс, который происходит по механизму про-ферментно-ферментного преобразования. С одной стороны, это обеспечивает нарастание сигнала: активация одной молекулы предшествующего уровня в системе свертывания приводит к активации от нескольких десятков до нескольких сотен тысяч последующих молекул (рис. 36). С другой стороны, многостадийность позволяет более гибко регулировать процесс.
В классическом каскаде свертывания крови выделяют 2 пути активации процесса:
| Рис. 36. Каскадный принцип усиления сигнала.Каждый предыдущий компонент системы свертывания активирует много последующих |
• Активация тканевым фактором (ТФ). Так как
ТФ не относится к плазменным факторам и
контактирует с кровью только при повреж
дении сосуда, то активация с его участием
обозначается как внешний путь свертывания.
• Активация ф.ХII при контакте с отрицатель
но заряженной поверхностью твердого тела,
или контактная активация. Поскольку фак
тор XII в норме присутствует в плазме, акти
вация с его участием обозначается как внут-
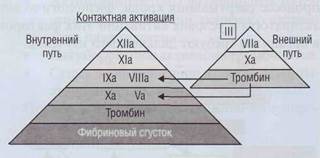
ренний путь свертывания (все факторы присутствуют в плазме в норме). Внешний и внутренний пути взаимодействуют между собой, а их разделение достаточно условно. Внешний и внутренний пути сходятся на факторе X. Последний со своим кофактором ф.Vа образует протромбиназу - ферментативный комплекс, который активирует протромбин с образованием тромбина. Образовавшийся тромбин поступает в ток крови и активирует фибриноген до фибрин-мономеров. Последние спонтанно соединяются, образуя полимеры фибрина. Условно свертывание плазмы (крови) делится на 2 основные фазы:
1) многоступенчатый этап, приводящий к акти
вации протромбина и превращению его в ак
тивный фермент - тромбин;
| Рис. 37. Каскад активации плазменного гемостаза |
2) конечный этап, в котором под влиянием тром
бина из фибриногена образуется фибрин.
Схема коагуляционного каскада плазменно
го гемостаза представлена на рис. 37.
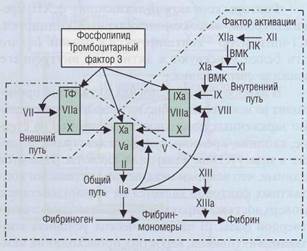
Внешний путь образования протромбиназы
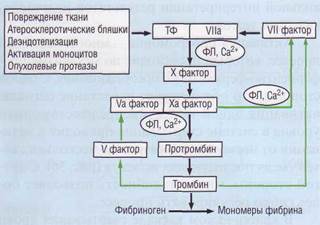
Внешний путь образования протромбиназы короткий, что ведет к быстрому образованию тромбина.
При контакте ТФ и ф.VIIа формируется комплекс, который активирует ф.Х. Фактор Ха при участии фактора Va, в присутствии ионов Са2+, на отрицательно заряженной фосфолипидной поверхно-
сти формирует протромбиназу. В настоящее время полагают, что внешний путь - основной физиологический путь запуска процесса свертывания крови. Подробно его значение будет описано в разделе «Современная теория свертывания крови».
Активность внешнего пути поддерживается за счет механизма положительной обратной свя-
Плазменные белки гемостаза
|
зи (рис. 38). Положительная обратная связь включается на нескольких этапах каскада свертывания. Наиболее существенными являются активация тромбином факторов VII и V.
Рис. 38. Внешний каскад свертывания крови. Начинается с контакта крови с тканевым фактором (ТФ), который, взаимодействуя с ф.VIIа, образует комплекс, активирующий ф, Х, Усиление активности внешнего каскада свертывания крови обеспечивается 2 механизмами положительной обратной связи
Внутренний путь образования протромбиназы. Факторы контактной активации
 Внутренний путь активации свертывания начинается с активации контактных факторов коа-гуляционного каскада: ф.ХII, прекалликреина и высокомолекулярного кининогена.
Внутренний путь активации свертывания начинается с активации контактных факторов коа-гуляционного каскада: ф.ХII, прекалликреина и высокомолекулярного кининогена.
Факторы контактной активации - ф.ХII, пре-калликреин, высокомолекулярный кининоген, С1-ингибитор - синтезируются в печени. In vitro эти белки участвуют в активации внутреннего каскада свертывания.
В лабораторных условиях активация происходит на некоторых небиологических отрицательно заряженных поверхностях, например на стекле, каолине, кремнии, сульфате декстрана, а также в присутствии эллаговой кислоты. Имеются данные, что важным механизмом активации контактных факторов является их взаимодействие с поверхностью, характеризующейся свойствами твердой фазы. В патологических условиях контактная активация, вероятно, происходит на мембранах клеток крови и эндотелия, а также при контакте с коллагеном субэндотелия.
Схематично взаимодействие белков при контактной активации показано на рис. 39. Видимо, в «подходящих условиях» происходит аутоактивация и взаимоактивация ф.ХII, ПК до активных ферментов. In vitro активация контактной системы приводит к активации ф.ХI, который в свою очередь активирует ф.IХ, образующий с ф.VIII теназный комплекс. Теназный комплекс (название комплекса происходит от английского слова ten - десять) активирует ф.Х, а далее процесс свертывания идет по уже описан-
ному пути. Поскольку сборка теназного комплекса происходит на фосфолипидной поверхности, для нее необходимо присутствие ионов кальция. Контактная фаза активации поддерживается положительной обратной связью. Тромбин активирует ф.VIII и -XI.
Физиологическое значение контактной активации, роль факторов контактной активации в процессе свертывания крови, физиологические активаторы и условия активации этих факторов в организме требуют дальнейшего изучения.
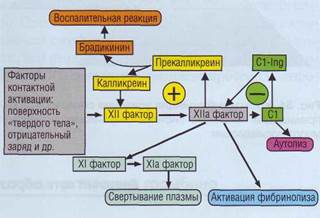
Рис. 39. Контактная фаза активации плазменных факторов. Контакт с поверхностью твердого тела вызывает активацию фактора XII, который запускает каскад свертывания плазмы, каскад активации фибринолиза, активацию калликреин-кининовой системы (положительная обратная связь) и активацию системы комплемента (отрицательная обратная связь)
Плазменные белки гемостаза
Классическая теория свертывания крови оставляла слишком много вопросов и противоречила клиническим данным. Например, с одной стороны, было неясно, какая поверхность в физиологических условиях является активатором, с другой стороны, почему дефицит факторов внутреннего пути (ф.VIII, -IX, -XI) приводит к выраженной кровоточивости при нормальной активности факторов внешнего пути, а глубокий дефицит факторов контактной активации, как правило, не сопровождается геморрагическим синдромом. В современной каскадно-матричной теории гемостаза эти противоречия разрешены.
С современной точки зрения, контактная активация играет большую роль во взаимодействии системы свертывания крови с другими протеоли-тическими системами крови (фибринолитичес-кой, ангиотензин-рениновой, калликреин-кини-новой, системой комплемента и др.).
В настоящее время изучены следующие функции белков контактной активации:
1. Брадикинин стимулирует повышение внутри
клеточной концентрации цАМФ и приводит к:
• Вазодилатации и снижению артериально
го давления.
• Активации системы фибринолиза путем
стимуляции секреции тканевого актива
тора плазминогена.
• Ингибированию активации тромбоцитов.
• Стимуляции репарации и росту гладкомы-
шечной ткани в поврежденных сосудах.
2. Прямое ингибирование тромбин-индуциро-
ванной активации тромбоцитов.
3. Активация фибринолиза.
• Непосредственная активация плазмино
гена калликреином и ф.ХIIа. Однако оба
этих белка значительно менее активны,
чем тканевой активатор и урокиназа.
• Активация калликреином проурокиназы
до активной сериновой протеазы - двух-
цепочечной урокиназы.
4. Блокада клеточной адгезии.
5. Антиангиогенное действие.
6. По-видимому, контактная активация играет
важную роль в активации свертывания кро
ви при взаимодействии крови с нефизиоло
гическими поверхностями, в частности при
установке искусственных протезов или искус
ственных клапанов сердца.
Внутренний путь образования протромбиназы (рис. 40) включает активирующее действие ф.ХПа на ф.Х1, который в свою очередь активирует ф.1Х. Поскольку значение контактной активации в процессе свертывания крови переосмыслено, физиологическая роль ф.Х1 изучается. Видимо, в физиологических условиях ф.Х1 в основном активируется тромбином. ф.Х1 довольно устойчив к инактивации ингибиторами и имеет длительный период полувыведения. Образовавшись в достаточном количестве, ф.Х1 увеличивает количество активного ф.1Х, за счет чего соответственно значительно возрастает концентрация тромбина, который в свою очередь активирует по механизму положительной обратной связи ф.1Х, -VIII и -V. В то же время избыток тромбина тормозит начало процесса фибри-
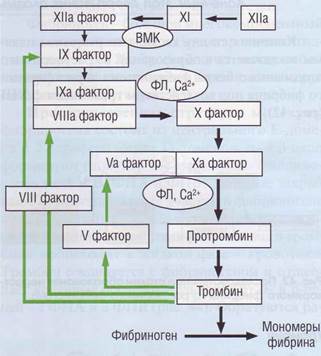
Рис. 40. Внутренний каскад активации плазменного гемостаза. Начинается с взаимной активации контактных факторов системы гемостаза, Фактор XIIа переводит фактор XI в ХIа. Фактор ХIа активирует фактор IX. Все последующие этапы активации свертывания по внутреннему пути требуют ионов Са2+ и зависят от присутствия фосфоли-пидов. Фактор IХа активирует фактор X, но эта реакция не очень эффективная. Однако появившийся тромбин активирует фактор VIII. Активный фактор Villa вместе с фактором IХа, ионами Са2+ и фосфолипидами очень эффективно активирует фактор Ха, Обратная связь поддерживает развитие процесса за счет активации тромбином ф.VIII, -IX и -V
Плазменные белки гемостаза

|
нолиза за счет активируемого тромбином ингибитора фибринолиза (TAFI).
Ингибитор С1-компонента комплемента (С1-ингибитор) является элементом системы контактной активации. Помимо комплемента, он ингибирует ф.ХIIа (см. раздел «Ингибиторы системы свертывания крови»).
Другим ингибитором процесса контактной активации в физиологических условиях является апротинин.
Рис. 41. Теназный и протромбиназный комплексы. Образование этих комплексов сопровождается резким увеличением активации соответственно фактора X и протромбина (фактор II)
|
|