Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника

Основные классы алкилирующих агентов
|
|
| Класс | Представитель | Структурная формула |
| Иприты сернистые | Иприт | S(CH2CH2Cl)2 |
| азотистые | Азотистый иприт | HN(CH2CH2Cl)2 |
| Эпоксиды | Этиленоксид | 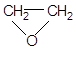
|
| Этиленимины | Этиленимин Триэтиленмеламин | 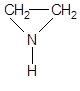 
|
| Алкилалкансульфонаты | Этилметансульфонат | C2H5OSO2CH3 |
| Метилметансульфонат | CH3OSO2CH3 | |
| b-Лактоны | b-Пропиолактон | 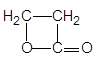
|
| Диазосоединения | Диазометан | CH2N ═ N |
| Нитрозосоединения | N-Нитрозо-N-метилуретан Диэтилнитрозамин N-Метил-N-нитро-N-нитрозогуанидин | 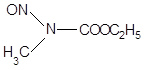
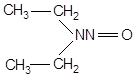 CH3N(NO)C(=NH)NHNO2
CH3N(NO)C(=NH)NHNO2
|
Алкилирование является одной из возможных химических модификаций нуклеотидов ДНК in vivoпод действием экзогенных мутагенов. Число известных химических веществ, способных вызывать модификации нуклеотидов ДНК по другим механизмам, быстро возрастает с расширением исследований в этой области. Среди таких мутагенов следует упомянуть азотистую кислоту, которая образуется из нитритов (NaNO2 и KNO2) в водных растворах при низких значениях pH. Азотистая кислота дезаминирует гуанин до ксантина, аденин до гипоксантина, а цитозин до урацила. В ДНК спаривание урацила с аденином приводит к транзициям GC®AT, гипоксантин вызывает обратную транзицию AT®GC, ксантин же не спаривается ни с одним из пиримидинов ДНК, и его включение оказывается летальным для клетки. Мутагенным действием обладают различные органические перекиси. Сама перекись водорода не оказывает мутагенного эффекта, но становится сильным мутагеном в сочетании с формальдегидом или ацетоном, у которых индуцирует образование свободных радикалов. Азид натрия – мощный ингибитор дыхания, также в ряде случаев обладает мутагенным действием, что связывают с накоплением в процессе метаболизма мутагенных перекисей. Перекиси индуцируют мутации и разрывы хромосом, имитируя мутагенное действие рентгеновских лучей, которые индуцируют образование различных реакционноспособных радикалов. И, наконец, среди химических мутагенов необходимо упомянуть аналоги нуклеозидов и оснований: 5-бромдезоксиуридин и 2-аминопурин, являющиеся сильными мутагенами.Спаривание с аденином 5-бромдезоксиуридина, обычно включающегося в ДНК вместо цитозина, приводит к образованию транзиций GC®AT. Обратные транзиции AT®GC возникают под действием 2-аминопурина.
Красители, обладающие способностью интеркалировать между основаниями ДНК, вызывают мутации со сдвигом рамки считывания. К таким красителям, в частности относятся хорошо известные бромистый этидий и производные акридина.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Промутагены, проканцерогены и их метаболическая активация. Рассмотренные выше химические соединения изначально обладают мутагенной активностью и для ее проявления не требуют каких-либо дополнительных модификаций. Однако существует множество химических соединений, которые сами по себе не проявляют мутагенную активность, но приобретают ее после серии биохимических превращений внутри организма под действием его ферментных систем. Такие химические соединения получили название промутагенов, а процесс их ферментативного превращения в мутагены – метаболической активации.
Любой организм в течение жизни подвергается непрерывной атаке экзогенными чужеродными химическими соединениями – ксенобиотиками, для защиты от накопления которых у него имеются эффективные ферментативные механизмы. Процесс метаболизма ксенобиотиков в организме традиционно разделяют на две фазы. Поскольку многие опасные для здоровья ксенобиотики гидрофобны и имеют тенденцию к накоплению в липидах клеточных мембран, обе фазы их внутриклеточного метаболизма связаны, прежде всего, с введением в их молекулы гидрофильных химических групп. Последующее повышение растворимости этих соединений в биологических жидкостях облегчает их выведение из организма. Таким образом, во время первой фазы метаболизма ксенобиотиков происходит ферментативное введение в их молекулы небольших полярных групп (например гидроксильных), что делает молекулы ксенобиотиков более водорастворимыми. С другой стороны, такие реакции подготавливают молекулы ксенобиотиков для вступления во вторую фазу метаболизма, во время которой происходит конъюгация преобразованных в первой фазе молекул ксенобиотиков с еще более полярными химическими группировками, в частности с остатками глюкуроновой кислоты, сульфатов или глицина.
Среди ферментов, принимающих участие в первой фазе метаболизма ксенобиотиков, наиболее хорошо изученными являются цитохромы группы Р-450. Эти ферменты в организме представлены большим числом (> 20) изоформ, каждая из которых, как правило, обладает широкой субстратной специфичностью, а все вместе они способны метаболизировать множество экзогенных и эндогенных химических соединений. Обобщенное уравнение химической реакции, осуществляемой цитохромами Р-450, представлено ниже:
S + NAD(P)H + O2 ® SO +NAD(P)+ + H2O,
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
где S – молекула субстрата, а SO – ее окисленная форма. Таким образом, для окисления субстрата цитохром Р-450 чаще всего использует электроны свободного кислорода, поступление которых к ферменту опосредовано NAD(P)H и флавопротеиновой редуктазой. Молекулы цитохрома Р-450 интегрированы в мембраны и при гомогенизации клеток ассоциированы с фракцией микросом. На рис. I.53 представлен механизм метаболической активации 1, 2-бензпирена (БП) – одного из наиболее сильных проканцерогенов. БП широко распространен в окружающей среде и в больших количествах образуется при горении нефтепродуктов, табака, а также при других термических воздействиях на органические соединения. Метаболические превращения БП по эпоксид-диольному пути начинаются с его эпоксидирования в положении 7, 8 определенной изоформой цитохрома Р-450, ассоциированной с мембранами микросом. Полученный эпоксид гидролизуется эпоксидгидролазой в соответствующий диол БП. Далее под действием другой изоформы цитохрома Р-450 происходит образование предпочтительно двух диастереомеров эпоксида в положении 9, 10 и других минорных производных (соединения в фигурных скобках, см. рис. I.53). Оказалось, что изомер (+)-анти-БПДЭ как in vitro, так и in vivo нестабилен и ковалентно связывается преимущественно с N2-атомом дезоксигуанозина ДНК своим С10-атомом с образованием аддуктов БПДЭ-ДНК.
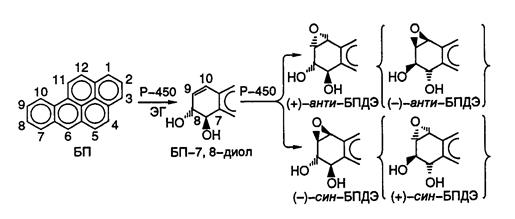
Рис. I.53. Механизм метаболической активации 1, 2-бензопирена (БП) микросомами животных
БПДЭ – 9, 10-эпоксид-1, 2-бензопирен-7, 8-дигидродиола, Р-450 – цитохром Р-450, ЭГ – эпоксидгидролаза
Механизм метаболической активации БП подробно рассмотрен здесь для иллюстрации общих принципов химического мутагенеза. Метаболическая активация ксенобиотиков с образованием реакционноспособных метаболитов является одной из основных причин генотоксичности множества химических соединений окружающей среды, с которыми организм контактирует на протяжении всей жизни. Список химических мутагенов экзогенного происхождения далеко не исчерпывается перечисленными химическими соединениями. По мере углубления исследований число известных мутагенов непрерывно растет благодаря созданию высокочувствительных методов оценки мутагенной активности. Известно, что почти все мутагены одновременно являются канцерогенами – веществами, вызывающими возникновение злокачественных опухолей, хотя обратное утверждение не всегда верно. Все это объясняет необходимость сохранения чистоты среды обитания человека, животных и растений.
Эндогенные мутагены. Выше уже упоминалось, что молекулы ДНК часто претерпевают in vivoтепловую депуринизацию, которая может быть спонтанным внутренним источником измененных нуклеотидов. Другим источником эндогенных мутаций служит самопроизвольное дезаминирование цитозина в составе ДНК с образованием урацила. 5-Метилцитозин – одно из модифицированных оснований ДНК, представляет собой " горячую точку " возникновения мутаций путем спонтанного дезаминирования, так как в результате удаления его аминогруппы образуется нормальное основание T, не распознаваемое системами репарации как мутантное.
К сожалению, не только окружающая среда может быть источником генотоксических химических соединений, отравляющих жизнь организма и нарушающих функционирование его генома. Свободные радикалы, которые образуются в организме во время нормального метаболизма, могут быть дополнительной и существенной причиной спонтанных мутаций. В частности, частота повреждений нуклеотидов ДНК, появляющихся под действием свободных радикалов кислорода, приближается к частоте мутаций, возникающих при депуринизации ДНК. В клетках свободные радикалы кислорода возникают в реакциях восстановления, в результате которых появляются чрезвычайно реакционноспособные промежуточные соединения. Наибольшую опасность для ДНК представляют радикалы гидроксила, супероксид и синглетный кислород, которые образуются в процессе дыхания, фагоцитоза и при повреждении клеток. Путем измерения содержания в моче человека 8-гидроксидезоксигуанозина и тимингликоля – основных модифицированных нуклеозидов и оснований, образующихся под действием радикалов кислорода, было установлено, что ежедневно в каждой клетке человека возникает ~10000 таких модифицированных нуклеотидов, мутагенный эффект которых в настоящее время доказан.
Другим источником эндогенных мутагенов в организме является метаболизм нормальной микрофлоры. Некоторые промежуточные соединения, возникающие при метаболизме аминокислот, желчных кислот и холестерина под действием бактерий организма человека и обладающие мутагенной и канцерогенной активностью, представлены в табл. I.19. Промоторами в канцерогенезе называют вещества, не имеющие канцерогенной активности, но ускоряющие процесс канцерогенеза под действием химических веществ – истинных канцерогенов. Как видно из таблицы, достаточно широк список веществ, обладающих всеми вышеупомянутыми активностями и образующихся под действием нормальной микрофлоры человека, обитающей, в частности в его кишечнике и на слизистых оболочках. Помимо этого в результате метаболизма в бактериальных клетках происходит активация нитратов с образованием мутагенных и канцерогенных N-нитрозаминов в кишечнике, желудке и ротовой полости человека. Описан также гидролиз обезвреженных ферментами детоксикации конъюгатов истинных канцерогенов.
Таким образом, многочисленные мутагены экзогенного и эндогенного происхождения создают вокруг и внутри любого многоклеточного организма тот мутагенный фон, к которому он вынужден приспосабливаться в процессе эволюции. Мутагенный фон " среды обитания" информационных макромолекул и, прежде всего, ДНК уникален для каждого вида живых организмов в силу особенностей их метаболизма и условий жизни. В соответствии с этим каждый биологический вид для того чтобы выжить, т.е. понизить частоту мутаций в конкретных генах до приемлемого уровня, обладает системами защиты от мутаций определенной эффективности. О механизмах функционирования таких систем защиты генетической информации речь пойдет ниже. А пока рассмотрим некоторые методы определения мутагенной активности химических соединений, применяемые для мониторинга окружающей среды.
Таблица I.19
|
|