Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Для новых пользователей первый месяц бесплатно. Чат-бот для мастеров и специалистов, который упрощает ведение записей: — Сам записывает клиентов и напоминает им о визите;
— Персонализирует скидки, чаевые, кэшбэк и предоплаты;
— Увеличивает доходимость и помогает больше зарабатывать; Начать пользоваться сервисом

Мегаспорогенез и развитие женского гаметофита
|
|
Наиболее важную роль в семязачатке играет нуцеллус, или мегаспорангий, в котором образуются мегаспоры и впоследствии развивается женский гаметофит.
Как уже было сказано, еще на ранней стадии развития семязачатка в нуцеллусе обособляется единственная археспориальная клетка (очень редко возникают 2 и более клеток), непосредственно превращающаяся в мегаспо-роцит. По размерам он больше остальных клеток нуцеллуса, имеет плотную цитоплазму и крупное диплоидное ядро. В результате редукционного деления ядра мегаспороцита возникают гаплоидные мегаспоры, составляющие линейную тетраду (рис. 244 В, 1, 3). Из самой нижней, сильно разрастающейся мегаспоры, в дальнейшем развивается женский гаметофит, а остальные мегаспоры отмирают (рис. 245 А).
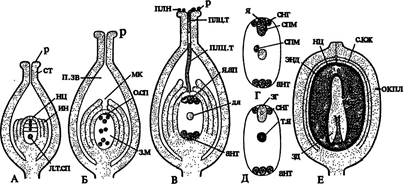
| Рис. 245. Развитие зародышевого мешка, оплодотворение, строение односеменного плода: А - тетрада мегаспор; Б - 8-ядерный зародышевый мешок; В - дифференциация клеток внутри зародышевого мешка; Г - двойное оплодотворение; Д - образование зиготы и вторичного ядра зародышевого мешка; Е - односеменной плод в продольном разрезе. |
Обозначения: ант — антиподы, д.я — диплоидное ядро, зг — зигота, зд — зародыш, з.м — зародышевый мешок, ин — интегументы, л.т.сп — линейная тетрада мегаспор, мк — микропиле, нц — нуцеллус, окпл — околоплодник, о.сп — 3 отмершие мегаспоры, п.зв — полость завязи, плн — пылинки, плц.т — пыльцевая трубка, р — рыльце, с.кж — семенная кожура, снг- синергиды, спм — спермий, ст — столбик, т.я — триплоидное ядро, энд — эндосперм, я — яйцеклетка, я.ап — ядерный аппарат
Женский гаметофит покрытосеменных называют зародышевым мешком. Он формируется следующим образом. Ядро увеличившейся в размерах мегаспоры митотически делится, возникшие 2 ядра расходятся к разным полюсам клетки. В ее центральной части формируется крупная вакуоль. При втором делении образуются 4 ядра, при третьем - восемь. К началу клеточной дифференциации зародышевого мешка исходная клетка достигает значительных размеров (рис. 245 Б). Параллельно с ее ростом разрастается и весь семязачаток, нуцеллус, как правило, истончается, а иногда, как у сложноцветных, исчезает полностью, так как содержащиеся в нем питательные вещества расходуются на развитие заростка. От каждого полюса клетки в ее центральную часть отходит по одному ядру, которые впоследствии сливаются, образуя диплоидное ядро центральной клетки зародышевого мешка.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
На микропилярном полюсе зародышевого мешка вокруг каждого из 3 оставшихся ядер обособляется цитоплазма, формируются оболочки и образуется яйцевой аппарат, состоящий из яйцеклетки и двух вспомогательных клеток — синергид. Яйцеклетка обычно крупнее, ее ядро находится в нижней части, под вакуолью, а в синергидах, наоборот, ядра располагаются в верхних частях клеток, над вакуолями. На халазальном полюсе зародышевого мешка таким же путем возникают 3 антиподы (рис. 245 В, Г).
Описанный, наиболее часто встречающийся 8-ядерный зародышевый мешок называют моноспорическим. Он образуется из одной (самой внутренней) мегаспоры тетрады. Однако 8-ядерные зародышевые мешки могут формироваться и другими способами (рис.246). У лука после 1-го деления мейоза возникает диада клеток. Одна клетка дегенерирует. Ядро второй клетки делится митотически, образующиеся ядра расходятся к разным полюсам клетки. Каждое из этих ядер соответствует ядру одной из 2 мегаспор диады. Формирующийся из двуядерной клетки зародышевый мешок называют биспорическим. У адоксы мегаспорогенез вообще не сопровождается цитокинезом, и каждое из 4 образующихся ядер соответствует ядрам неразвившейся тетрады мегаспор. Возникший из такой 4-ядерной клетки зародышевый мешок называют тетраспорическим.
Эти примеры показывают, что у покрытосеменных растений существуют разные механизмы, ускоряющие развитие женского гаметофита. Другие отклонения в строении зародышевого мешка связаны с уменьшением или увеличением числа клеток и числа ядер, их плоидности. Особенно сильно варьируют числа антипод.
Таким образом, женский гаметофит покрытосеменных по сравнению с женскими гаметофитами всех других разноспоровых растений достиг крайней степени редукции. Он состоит из крупной диплоидной центральной клетки, окруженной оболочкой мегаспоры, и 6 находящихся внутри нее мелких клеток. Женский гаметофит покрытосеменных растений утратил архегонии, сохранив способность к формированию женской гаметы - яйцеклетки.
Естественно, возникает вопрос о возможности гомологизации зародышевого мешка с женскими заростками других разноспоровых растений. Все многочисленные точки зрения практически могут быть сведены к 3 основным.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
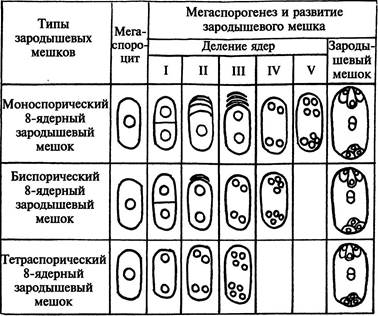
| Рис. 246. Разные типы формирования 8-ядерных зародышевых мешков |
1. Зародышевый мешок - типичный гаметофит, состоящий из вегетативной части (синергид и антипод) и редуцированного архегония, от которого сохранилась одна яйцеклетка.
2. Зародышевый мешок имеет 2 редуцированных архегония, расположенных в микропилярной и халазальной его частях. При этом яйцеклетку считают гомологом одной из антипод. 2 синергиды и 2 антиподы рассматривают как шейковые канальцевые клетки, а полярные ядра - как ядра брюшных канальцевых клеток. Однако в настоящее время выяснено, что синергиды представляют собой сестринские клетки, а ядро яйцеклетки - сестринское одному из полярных ядер. Таким образом, эта гипотеза оказалась несостоятельной.
3. Зародышевый мешок - гомолог женского гаметофита остальных разноспоровых растений, так как возникает из мегаспоры, но он настолько видоизменился в процессе эволюции, что нецелесообразно пытаться найти гомологию между его отдельными структурами.
8-ядерный зародышевый мешок соответствует свободно-ядерной стадии развития женского гаметофита голосеменных и некоторых разноспоровых папоротников, например, сальвинии. При общности начальных стадий развития всех женских гаметофитов, конечные стадии развития зародышевого мешка сильно изменились и приобрели специфические особенности, не имеющие гомологов среди голосеменных. В связи с этим, формирование зародышевого мешка можно рассматривать как следствие эволюционного ускорения, сопровождающегося утратой способности к образованию типичных архегониев и вегетативной части заростка. Кстати, этот процесс, хотя и в другом варианте, происходил и у гнетовых из отдела голосеменных, у которых отмечены все стадии развития женских заростков - от нормально развитого эндосперма с архегониями у эфедры до своеобразных гаметофитов, не имеющих архегониев, у гнетума и вельвичии.
|
|