Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Предки и географическое происхождение современного домашнего скота
|
|
Одним из основных направлений исследований, затрагивающих сферы деятельности археологической и генетической наук, является определение очагов одомашнивания современных домашних животных (Zeder и др., 2006). При этом, совместные исследования открывают новые факты, связанные с географией происхождения и разнообразием домашнего скота. В настоящее время известно, что практически все основные виды домашних животных явились продуктом нескольких событий одомашнивания в различных географических зонах (табл. 4 и рис. 3), а также что последующая за начальными событиями одомашнивания генетическая интрогрессия между дикими родственными формами и их домашними аналогами встречалась часто.
Следует отметить, что представляющиеся независимыми события одомашнивания, в ряде случаев не являются исторически совершенно независимыми. По-видимому, в некоторых случаях могло происходить перемещение нескольких одомашненных особей из одной области в другую и последующее поглощение их генетического материала местной дикой фауной (Zeder и др., 2006). И наоборот, древние следы одомашнивания в данной зоне могут быть скрыты вследствие более позднего завоза домашнего скота в эту зону из других центров одомашнивания животных. Остеометрическая информация, полученная в археологических исследованиях, и изучение ДНК ископаемого домашнего скота могут послужить полезными инструментами для разрешения такого рода вопросов.
Приручение домашнего скота, как это принято сейчас считать, произошло в 12 центрах мира (рис. 3).
Характерно, что не все центры одомашнивания животных тесно связаны с родиной наших культурных видов растений (см. рис. 2). В то время, как в некоторых случаях (например, в зоне Плодородного полумесяца), центры одомашнивания как растений, так и животных объединены, в других (к примеру, на африканском континенте) – окультуривание растений и одомашнивание животных, скорее всего, происходили независимо друг от друга. Хотя для некоторых
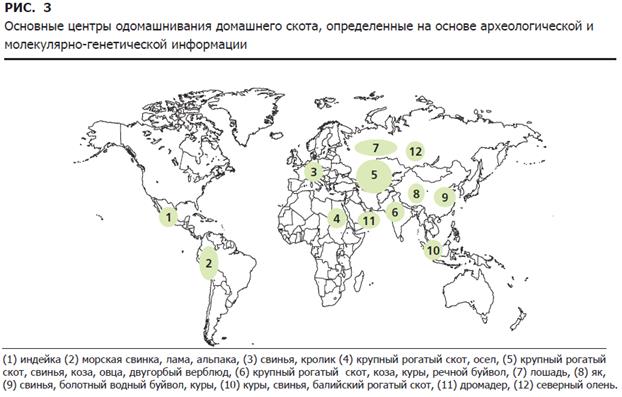
видов животных еще нельзя с уверенностью указать центры их одомашнивания, для других – такие географические зоны можно указать с определенной достоверностью. К ним относятся: горная цепь Анд Южной Америки (ламы, альпака, морские свинки); Центральная Америка (индейки, мускусные утки); северо-восточная Африка (крупный рогатый скот, ослы); юго-западная Азия, включая Плодородный полумесяц (крупный рогатый скот, овцы, козы, свиньи); долина реки Инд (крупный рогатый скот, козы, куры, речные буйволы); Юго-Восточная Азия (куры, балийский скот); восточный Китай (свиньи, куры, болотные буйволы); гималайское плато (яки); Северная Азия (северный олень). Вполне вероятно, что зарождение дромадера произошло в южной части Аравийского полуострова, бактриана – в области, занимаемой ныне Исламской республикой Иран, а лошади – в европейских степях.
Одомашнивание животных происходило не только в разных зонах, но и в разное время. Однако до сих пор идут дискуссии о времени одомашнивания разных видов животных. Основной проблемой при этом является то, что одомашниваемые животные не могли по морфологическим характеристикам отличаться от своих диких предков, а, следовательно, результаты исследований, основанных на использовании морфологических маркеров, могут дать заниженную оценку древности одомашнивания (Dobney, Larson, 2006). Кроме того, современные молекулярные методы оценки времени, хотя и не зависят от морфологических изменений, еще несовершенны и имеют большие ошибки при определении временных шкал.
Вместе с тем, существуют новые методы, включая метод демографического профиля, и методы калибровки молекулярных часов, которые могут быть использованы для идентификации более точного времени одомашнивания животных (Zeder и др., 2006).
Новые результаты археологических и генетических исследований постоянно улучшают наши знания о происхождения видов домашнего скота. Известно, что первым видом одомашненных животных была собака. Это, вероятно, произошло около 14 000 лет назад; уже в то время собаки использовались для охоты и охраны. Однако, до сих пор не ясно, в каком месте происходило начальное одомашнивание. Поскольку среди современных собак обнаруживается много материнских линий, значит интрогрессия с их диким предком – волком (Canis lupus) – обитавшем в Старом Свете осуществлялась много раз. Домашние собаки в Новом Свете, скорее всего, имеют европейское происхождение, о чем свидетельствуют результаты анализа митохондриальных линий (Wayne и др., 2006).
Козы были одомашнены уже 10 000 лет назад в Загросе, район Плодородного полумесяца (Zeder, Hesse, 2000). Безоаровый козел (Capra aegagrus) был, вероятно, одним из предков домашней козы, однако, возможно и другие виды, как например, C. falconeri, внесли свой вклад в формирование современного генофонда. В настоящее время у домашних коз идентифицированы пять различных материнских митохондриальных последовательностей (Luikart и др., 2001; Sultana и др., 2003; Joshi и др., 2004). Из них одна преобладает численно и присутствует во всех популяциях, вторая, - скорее всего, имеет современное происхождение. Все они, вероятно, отражают изначальный процесс приручения коз в Плодородном полумесяце, где по археологическим данным определяют две или три зоны одомашнивания (Загрос, Таврские горы, долина Иордана). Другие митохондриальные линии коз географически долее ограничены и, возможно, соответствуют дополнительным центрам одомашнивания или интрогрессии, включая долину Инда (Ferná ndez и др., 2006).
Овцы также, вероятно, были одомашнены в зоне Плодородного полумесяца, приблизительно 8-9 тыс. лет назад. Археологическая экспертиза определяет два независимых очага одомашнивания овец в Турции: верхняя долина Евфрата в восточной Турции, и центральная Анатолия (Peters и др., 1999). Три вида диких овец (уриал, Ovis vignei; аргали, O. ammon.; и евразийский муфлон, O. musimon/orientalis), идентифицированы как предки современных домашних овец (Ryder, 1984) или, по крайней мере, происходила их интрогрессия с некоторыми локальными породами. Однако недавние генетические исследования не подтвердили эти данные относительно уриала и аргали (Hiendleder и др., 1998), что послужило основанием считать азиатского муфлона (O.orientalis), широко распространенного в географической зоне от Турции до, по крайней, Исламской республики Иран, единственным прародителем домашней овцы. От одичавших овец, как сейчас полагают, ведет свое происхождение Европейский муфлон (O. musimon). У домашних овец зарегистрированы четыре основных линии митохондриальной ДНК (Hiendleder и др., 1998; Pedrosa и др., 2005; Tapio и др., 2006), из которых одна или две могут быть связаны с событиями одомашнивания, а остальные – с последующей интрогрессией с дикими видами. До сегодняшнего дня не существует никаких четких доказательств связи этих митохондриальных линий с фенотипическим разнообразием овец (в частности, с типами жирнохвостых, тощехвостых и курдючных овец).
Предком домашней свиньи считается дикий кабан (Sus scrofa). Полученные обширные зоогеографические данные свидетельствуют, что свиньи были одомашнены приблизительно 9 тыс. лет назад на Ближнем Востоке. Материал, обнаруженный в Восточной Анатолии, свидетельствует об одомашнивании свиньи и дальнейших постепенных изменениях, произошедших в морфологии и демографическом профиле животных за тысячи лет. Существуют также археологические и генетические доказательства существования другого независимого центра одомашнивания свиньи в Восточной Азии (Guiffra и др., 2000). Как минимум, еще 16 различных подвидов дикого кабана описаны в Евразии и Северной Африке, поэтому не удивительно, что проведенный недавний обзор разнообразия митохондриальных ДНК среди евразийских свиней и дикого кабана свидетельствует о сложной картине одомашнивания свиньи в, по крайней мере, пяти-шести центрах в рамках географического ареала дикого вида (Larson и др., 2005).
Одомашнивание крупного рогатого скота особенно хорошо изучено. Существует доказательство одомашнивания трех различных подвидов туров (Bos primigenius) в трех зонах. B. primigenius primigenius, одомашненный в зоне Плодородного полумесяца около 8 000 лет назад, и B. p. opisthonomous, возможно одомашненный еще 9 000 лет назад в северо-восточной части африканского континента (Wendorf, Schild, 1994), являются предками безгорбого скота B.taurus Ближнего Востока и Африки, соответственно. Горбатый скот зебу (Bos indicus), как полагают, был одомашнен позднее, приблизительно 7 000 - 8 000 лет назад, в долине реки Инд на территории современного Пакистана (Loftus и др., 1994; Bradley и др., 1996; Bradley, Magee, 2006). Недавно выдвинули предположение о существовании четвертого центра одомашнивания в Восточной Азии (Mannen и др., 2004), однако считается не ясным, произошло ли одомашнивание независимо или представляет интрогрессию местного тура со скотом Ближневосточного происхождения.
Предком домашнего индийского буйвола (Bubalus bubalus), несомненно, является дикий буйвол Азии. На основании исследований фенотипов, кариотипов и митохондриальной ДНК выделены два его основных типа (Tanaka и др., 1996): речной буйвол в Индостане, на Ближнем и Среднем Востоке и в Восточной Европе; и болотный буйвол в Китае и странах Юго-Восточной Азии. Оба типа скрещиваются в северо-восточной части Индостана. Они были, вероятно, одомашнены независимо. Предполагаемые центры одомашнивания речного буйвола находятся в долине Инда и/или в долинах Евфрата и Тигра, и процесс начался около 5 000 лет назад, а болотного буйвола - в Китае, где одомашнивание происходило примерно 4 000 лет назад вместе с началом культивирования здесь риса.
Существуют разные мнения о происхождении и одомашнивании лошади (Equus caballus). Как факт, можно считать, что ее дикий предок вымер. В качестве предполагаемых диких предков лошади рассматриваются два вида: тарпан (E. ferus) и лошадь Пржевальского (E. przewalskii). Лошадь Пржевальского, хотя и является близким родственником дикого предка, однако, скорее всего не является прямым прародителем домашних лошадей (Olsen и др., 2006; Vilà и др., 2006). В исследованиях трудно оценить, дикой или домашней лошади принадлежат археологические останки. Имеются доказательства, что лошадь была одомашнена приблизительно в 3700-3100 г. до н.э. (Медный век) на севере Казахстана (Ботайская культура) (Olsen, 2006). Однако результаты последних молекулярных исследований показывают, что вероятнее всего лошадь происходит из нескольких популяций различных географических областей. При этом, еще не установлено, произошел ли единственный случай одомашнивания и последующая интрогрессия, или имели место многократные независимые случаи одомашнивания (Vilà и др., 2001; Jansen и др, 2002).
Одомашнивание осла Equus asinus, напротив, выглядит более ясным. Митохондриальные исследования ДНК подтвердили африканское происхождение домашнего осла и исключили азиатского дикого осла как возможного его прародителя (Beja-Pereira и др., 2004). Две митохондриальные линии предполагают два события одомашнивания осла. Одна линия тесно связана с нубийским диким ослом (E. asinus africanus), который до сих пор существует в природе, обитая в северо-восточной части Судана вблизи Красного моря. Другая линия указывает на некоторую родственную связь современных животных с сомалийским диким ослом (E. asinus somaliensis), что свидетельствует также об их африканском происхождении, хотя возможность одомашнивания в соседних областях (Аравийский полуостров или Плодородный полумесяц) пока что не исключена. Археологические доказательства, полученные в Египте, подтверждают африканский центр одомашнивания осла, и предполагают, что одомашнивание происходило около 6 000 - 6 500 лет назад (Clutton-Brock, 1999).
Домашний як (Poephagus grunniens) является эндемиком Центральной Азии и хорошо приспособлен к холодным и высокогорным условия обитания. Пастбищное содержание яков широко распространено в Центральном азиатском высокогорье, и начало его использования было решающим событием, обеспечившим возможность круглогодичного проживания человека в высотных зонах Гималайского плоскогорья. Возможно, это послужило толчком к формированию тибетско-бирманского населения в этой области. Сегодня дикие яки (P. mutus) все еще обитают на Цинхай-Тибетском нагорье, но они, скорее всего, подверглись сильной интрогрессии с одичавшим домашним яком. Идентифицированы три митохондриальных последовательности ДНК. Однако сходное географическое распространение митохондриальной ДНК свидетельствует о единственном случае одомашнивания в восточной части Цинхай-Тибетском нагорья (Qi, 2004; Guo и др., 2006). Полученные молекулярные данные также указывают, что распространению домашних яков способствовали два различных миграционных маршрута из их центра одомашнивания: як достиг «памирского узла” по западному пути через Гималаи и горы Кунь-Лунь; и достиг территорий Монголии и современной Российской Федерации через Монгольскую Южную Гоби и алтайскую Гоби.
Как и в случае с яком, одомашнивание северного оленя (Rangifer tarandus) дало возможность кочевым общинам осваивать новые среды обитания, не подходящие для содержания других видов домашних животных. Об одомашнивании северного оленя известно очень мало. Дикий северный олень, вероятно, был последним крупным одомашненным представителем класса млекопитающих. Археологическими исследованиями установлено, что самым ранним центром одомашнивания северного оленя являлись Алтайские горы Сибири, и процесс одомашнивания начался около 2 500 лет назад. Это свидетельствует о том, что уже в то время северные олени использовались как транспортное средство (Skjenneberg, 1984). Пока нет достоверных данных о том, как процесс одомашнивания этого вида достиг Европы. Полагают, что он мог независимо протекать в Скандинавии или был заимствован народами Саамы (Saami) у других северных евразийских кочевых племен. Ориентировочно считают, что саамы начали заниматься оленеводством приблизительно в XVII веке. В Северной Америке дикий северный олень известен как карибу, полагают, что он никогда не был одомашнен на этом континенте (Clutton-Brock, 1999).
Предполагают, что одомашнивание верблюда бактриана (Camelus bactrianus) произошло в области современной Исламской республики Иран/ Туркмении, или, возможно, в южном Казахстане, северо-западной Монголии или Северном Китае (Bulliet, 1975; Peters, von den Driesch, 1997). Самый древний центр одомашнивания верблюдов бактрианов определен в месте Сахри-Сокта (Sahr-i Sokta) в центральной части Исламской республики Иран, где были обнаружены верблюжьи кости, экскременты, и сотканные волокна, датированные приблизительно 2600 годом до н.э. (Compagnoni, Tosi, 1978). Результаты последних генетических исследований показывают, что популяции диких верблюдов (C. ferus) пустыни Гоби, успешно скрещивающиеся с домашними видами, вероятно, не являются прямыми материнскими предками домашних или одичавших верблюдов (Jianlin, и др., 1999). Дикий предок одногорбого верблюда дромадера (C. dromedarius) считается в настоящее время вымершим. Предполагается, что одомашнивание верблюдов началось около 5 000 лет назад в юго-восточной части Аравийского полуострова.
Происхождение южноамериканских верблюдовых в настоящее время установлено. Гуанако (Lama guanicoe) и викунья (Vicugna vicugna) являются предковыми видами домашней ламы (Lama glama) и альпаки (Vicugna pacos), соответственно (Kadwell и др., 2001). Археолого-зоологические исследования указывают центральные перуанские Анды как центр доместикации альпаки (6 000 - 7 000 лет назад). Предполагают, что лама могла быть одомашнена в тот же самый период в Андах, в районе озера Титикака. При этом выявлено широкомасштабные интрогрессии между этими двумя домашними видами (Wheeler и др., 2006). Этот продолжающийся процесс гибридизации, начался, вероятно, в эпоху испанской колонизации, которая разрушила традиционные структуры разведения этих видов.
Предком балийского рогатого скота является бантенг (Bos javanicus), три подвида которого признаны исчезающими. Характерно, что нет никаких свидетельств существования дикого предка этого вида животных на острове Бали. Балийский скот мог быть одомашнен на острове Ява и/или на Индокитайском полуострове. Обнаружена интрогрессия B. taurus и B. indicus в балийский скот, а генетический материал балийского скота обнаружен в некоторых юго-восточных азиатских породах крупного рогатого скота, что свидетельствует о его более широком распространении в прошлом (Felius, 1995).
Предок митхуна (mithun) (B. frontalis) – гаур (B. gaurus). Как и в случае с балийским рогатым скотом, центр его одомашнивания неизвестен. Археологические раскопки в северо-восточном Таиланде (Non Nok Tha) свидетельствуют, что оба вида были одомашнены уже 7 000 лет назад (Higham (1975), цитируется по Felius, 1995).
Домашние куры (Gallus domesticus) происходят от дикой красной джунглевой курицы (Gallus gallus), пяти возможных ее подвидах. Проведенные ранее молекулярные исследования выявили единственный центр происхождения - в Юго-Восточной Азии (Таиланд) (Fumihito и др., 1994; 1996). Однако последующие исследования идентифицировали, как минимум 6 генетических материнских линий (Liu и др., 2006), что предлагает несколько центров их одомашнивания. Археологические данные указывают на центры одомашнивания кур в районе долины Инда (5 000 лет назад) и в восточном Китае (возможно, 7 500 - 8 000 лет назад) (West, Zhou, 1988).
|
|