Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника

⚡️ Для новых пользователей первый месяц бесплатно. А далее 290 руб/мес, это в 3 раза дешевле аналогов. За эту цену доступен весь функционал: напоминание о визитах, чаевые, предоплаты, общение с клиентами, переносы записей и так далее.
✅ Уйма гибких настроек, которые помогут вам зарабатывать больше и забыть про чувство «что-то мне нужно было сделать».
Сомневаетесь? нажмите на текст, запустите чат-бота и убедитесь во всем сами!
Селективная защита генов от мутаций
|
|
Во всех предыдущих рассуждениях речь шла о глобальной защите функционально значимых участков гипотетического генома от спонтанных и индуцируемых мутаций некодирующими последовательностями нуклеотидов. При этом для простоты рассуждений предполагалось, что распределение нуклеотидов геномной ДНК и самих мутагенов в интерфазном ядре гомогенно. В реальном геноме эукариот распределение последовательностей нуклеотидов геномной ДНК в интерфазном ядре далеко не однородно. Достаточно вспомнить, что геном эукариот всегда представлен несколькими хромосомами (в частности диплоидный геном человека заключен в 46 хромосомах), ДНК каждой пары из которых обладает уникальными первичной и пространственной структурами. ДНК индивидуальных хромосом в интерфазном ядре компартментализована, а плотность упаковки ДНК в различных участках индивидуальных интерфазных хромосом неравномерна.
В этом отношении наиболее изучена и показательна хромомерная организация гигантских политенных хромосом, образующихся в клетках некоторых типов животных и растений. Для таких хромосом в интерфазном ядре характерны различные уровни компактизации хроматина вдоль хроматид, что морфологически проявляется в формировании визуально обнаруживаемых, поперечно расположенных дисков и междисков. В этих хромосомах выделяют три уровня компактизации ДНК. Дискам (неактивным районам политенных хромосом) свойственен максимальный 100–380-кратный уровень компактизации хроматина по отношению к свободной ДНК. С началом транскрипции, т.е. при переходе участков интерфазных хромосом в активное состояние, уровень компактизации понижается до ~40-кратного, и ДНК декомпактизуется еще сильнее в условиях максимальной транскрипции, когда молекулы РНК-полимеразы движутся вдоль ДНК одна за другой на небольшом расстоянии друг от друга, как это имеет место, например в кольцах Бальбиани. Уровень упаковки ДНК в хроматине метафазных хромосом самый высокий: исходная длина ДНК в этом случае уменьшается в 6000–7000 раз.
Следует подчеркнуть, что компактизация ДНК вдоль хроматид в интерфазных политенных хромосомах высоко упорядочена в индивидуальных хромосомах генома и обладает абсолютной видовой специфичностью. Это позволяет использовать рисунок поперечной исчерченности индивидуальных политенных хромосом для их идентификации и физического картирования генов. В настоящее время установлено, что некоторые большие гены Drosophila (например ген dunce, кодирующий фосфодиэстеразу циклического АМР, а также гены E74 и Shaker) локализованы в нескольких дисках и междисках и, следовательно, неравномерно компактизованы по своей длине. Различные уровни упаковки ДНК наблюдают в разных районах одних и тех же политенных хромосом, и для этих уровней, по-видимому, характерны более тонкие градации, чем те, которые выявляются при визуальных наблюдениях в виде поперечной исчерченности политенных хромосом.

Рис. I.64. Способы проникновения мутагенов в отдельные микрокомпартменты ядра (а), возможная роль интронов в дифференциальной защите генов (б) и расположение летальных генов на хромосоме 2 дрожжей (в)
а – мутагены, образующиеся вне микрокомпартмента, содержащего ДНК (слева) и внутри него (справа); б – гипотетический ген в виде " розетки" с интронами в виде петель, окружающих объединенные экзоны. Слева – структура гена с расправленными петлями, справа – их гипотетическая реальная пространственная структура, образующая защитную оболочку вокруг экзонов; в – расположение известных жизненно важных генов на хромосоме 2 без предварительной обработки исходных данных (1), а также после обработки с учетом кластеров и гипотетических летальных генов (2). Гены, расстояние между которыми < 10 т.п.о., объединяли в кластеры (подчеркнуты), которые на графике 2 изображали в виде отдельных точек с доверительными интервалами, лежащими между первым и последним нуклеотидами кластера. По мере необходимости вводили гипотетические гены (пропуски между точками на графике 2). Указаны коэффициент детерминированности R2, описывающий точность аппроксимации полученных данных к линейной функции, и ее уравнение
У большинства хромосом соматических клеток эукариот не происходит политенизации ДНК, и соответствующая часть геномной ДНК заключена в одной хроматиде. Тем не менее, несмотря на то что в интерфазных ядрах большинства соматических клеток визуально не выявляется четкая хромомерная структура индивидуальных хромосом, нет основания полагать, что их хроматиды организованы принципиально иначе, чем в политенных хромосомах. Действительно, у всех метафазных хромосом соматических клеток животных после их гистохимической окраски выявляется высокоспецифичная поперечная исчерченность (бэндинг), и этот рисунок может быть следствием компактизации высокоупорядоченных хромомерных структур, исходно присутствующих в интерфазных хромосомах. Если такое предположение верно, то у любых интерфазных хромосом соматических клеток эукариот расположение хромомеров вдоль хроматид, а, следовательно, и уровни компактизации ДНК видоспецифичны и являются постоянной характеристикой индивидуальных хромосом и биологических видов в целом.
Рассматривая индивидуальную интерфазную хромосому как потенциальную мишень для мутагенов, можно заключить, что защищенность генетической информации ее индивидуальных локусов от их действия будет прямо пропорциональна уровням компактизации заключенной в них ДНК, т.е. концентрации последовательностей нуклеотидов на единицу ядерного объема, занимаемого этими участками генома. Рассмотрим подробнее процессы, которые могут происходить в микрокомпартментах (отдельных хромомерах) хромосом при их контакте с химическими внутриядерными мутагенами (рис. I.64, а). Как и в случае целых ядер эукариотических клеток, химические мутагены поступают в микрокомпартменты двумя различными путями: образуясь непосредственно внутри них, например под действием ионизирующего излучения, и путем радиальной диффузии через поверхность микрокомпартментов (индивидуальных хромомеров). В первом случае частота мутаций, возникающих на участке заключенной в микрокомпартмент хромосомной ДНК в результате контактов с образующимися здесь же химическими мутагенами, будет обратно пропорциональна концентрации этой ДНК в микрокомпартменте, т.е. уровню ее компактизации в хромомере. Действительно, в продуктивных (приводящих к мутациям) взаимодействиях нуклеотидов ДНК с мутагенами участвует доля нуклеотидов ДНК, пропорциональная количеству нуклеотидов, контактирующих c мутагенами. Эта доля уменьшается при увеличении концентрации нуклеотидов-акцепторов мутагенов в микрокомпартменте. При условии взаимодействия всех k мутагенов с K нуклеотидами рассматриваемого микрокомпартмента (с последующим образованием аддуктов нуклеотид-мутаген и мутаций) частота мутаций F на данном участке ДНК будет равна:
F = 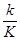 .
.
В более общем виде это выражение можно записать, как:
F = 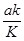 , (4),
, (4),
где a – коэффициент эффективности взаимодействия мутагенов с нуклеотидами-акцепторами (0 < a 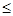 1). Если количество нуклеотидов и молекул мутагенов в индивидуальном микрокомпартменте выразить через их концентрации в нем (соответственно C и c), то выражение (4) примет вид:
1). Если количество нуклеотидов и молекул мутагенов в индивидуальном микрокомпартменте выразить через их концентрации в нем (соответственно C и c), то выражение (4) примет вид:
F =  ,
,
поскольку по определению k = cV, а K = CV (V – объем микрокомпартмента, занимаемого индивидуальным хромомером).
Если предположить, что стационарная концентрация мутагенов, образующихся внутри интерфазного ядра эукариотических клеток, одинакова во всех его частях, то частота мутаций, вызываемых такими агентами в индивидуальных хромомерах эукариотических хромосом, будет обратно пропорциональна концентрации нуклеотидов ДНК этих хромомеров в соответствующих микрокомпартментах, т.е. обратно пропорциональна плотности упаковки ДНК в хромомере. Следовательно, локальная плотность упаковки цепей ДНК в интерфазном ядре эукариотических клеток может быть важным фактором, определяющим частоту мутаций в соответствующих генетических локусах. При этом необходимо учитывать, что белки являются неотъемлемой частью хроматина эукариотических клеток, и они также могут оказывать влияние на частоту мутаций в индивидуальных хромомерах, затрудняя доступ мутагенов к нуклеотидам ДНК.
Судьба мутагенов, поступающих в ядро эукариотической клетки из цитоплазмы в результате радиальной диффузии, складывается иначе (см. рис. I.64, а, слева). По-видимому, значительная их часть взаимодействует с нуклеотидами ДНК, расположенной на периферии ядра или внутриядерных хромомеров, и степень защиты как ядра в целом, так и его отдельных микрокомпартментов прямо зависит от плотности такой защитной оболочки. Вторым фактором, регулирующим глубину проникновения мутагенов в ядро и его микрокомпартменты, является реакционноспособность самих мутагенов. Данный параметр, ранее обозначенный как коэффициент эффективности взаимодействия мутагенов с нуклеотидами a (уравнение (4)), определяет время жизни мутагена в ядре, т.е. как скоро молекула мутагена прореагирует с потенциальными нуклеотидами-акцепторами, которые мутаген встречает в процессе диффузии внутри ядра. Третьим фактором, от которого зависит эффективность защиты кодирующих жизненно важных последовательностей нуклеотидов генома от мутаций, вызываемых такими мутагенами, является пространственное расположение самих внутриядерных последовательностей. Очевидно, что вероятность встречи с мутагенами и взаимодействия с ними больше у нуклеотидов последовательностей, локализованных на периферии ядра или хроматиновых доменов – хромомеров. С учетом этого фактора можно полагать, что максимальная защита имела бы место в том случае, если бы мутагены первыми встречали избыточные некодирующие последовательности, мутации в которых нейтральны в отношении влияния на жизнеспособность клеток многоклеточного эукариотического организма.
В последнее время появляется все больше экспериментальных данных, свидетельствующих о высокоупорядоченном распределении специфических последовательностей нуклеотидов в интерфазных ядрах эукариот. Одна группа данных такого рода, связанных с упорядоченной конденсацией нитей ДНК вдоль хроматид индивидуальных интерфазных хромосом, была уже обсуждена выше. Помимо этого, активно рассматривается модель петельно-доменной организации генов в эукариотических хромосомах. Неслучайное распределение MAR- и SAR-последовательностей в геномной ДНК эукариот и их ассоциация с белками ядерного матрикса (скэффолда) в интерфазе клеточного цикла, по-видимому, весьма специфически контролируют пространственное расположение протяженных участков интерфазных хромосом в ядрах. Кроме того, на более низком (хромомерном) уровне компактизации интерфазных хромосом также обнаружены специфические способы укладки ДНК в виде больших и малых петель, вероятно, относящихся к хромомерам и " розеткам" соответственно (см. рис. I.64, б). Не исключена возможность, что в соответствии с моделью Н.А. Резника и соавторов (1991 г.) образование розеток из ДНП-петель специфически отражает интрон-экзонную структуру конкретных генов. Если такая модель соответствует действительности, то петли интронов генов, заключенных в розетки, могут создавать специфическую защитную оболочку, предохраняющую экзоны генов от контакта с мутагенами. В этой связи соотношение суммарных размеров интронов и экзонов индивидуальных генов может оказаться существенным параметром, характеризующим уровень защищенности функционально значимых участков генов от химического мутагенеза. Чем больше отношение суммарной длины интронов к суммарной длине экзонов в конкретном гене, тем более плотную защитную оболочку интроны могли бы сформировать вокруг компактизованных экзонов. Такое соотношение может указывать на уровень защищенности конкретных генов (или даже их частей) от химических мутагенов.
Описанные выше структурные особенности организации генетического материала позволяют предполагать разную и генетически детерминированную доступность индивидуальных участков интерфазных хромосом для химических мутагенов. Имеются, по крайней мере, три группы экспериментальных данных, указывающих на то, что различные участки одного и того же генома спонтанно изменяются с помощью мутаций с разной скоростью. Прежде всего, это прямые экспериментальные указания на неодинаковую мутабильность как различных генов одного и того же генотипа, так и одного и того же гена в генотипах разных биологических видов.
При другом подходе к исследованию этой проблемы путем компьютерного анализа баз данных последовательностей нуклеотидов было установлено, что степень эволюционной дивергированности конкретных участков генома у разных видов млекопитающих непостоянна и может изменяться от локуса к локусу. Скорость накопления замен нуклеотидов в конкретных локусах приблизительно одинакова как для кодирующих последовательностей нуклеотидов генов (экзонов), так и для соседствующих с ними некодирующих последовательностей, казалось бы, не подверженных давлению отбора. Этот, на первый взгляд, странный результат вполне естественно объясняется в рамках предлагаемой модели дифференциальной защиты различных хромосомных локусов от спонтанных мутаций как следствие упорядоченного гетерогенного распределения геномной ДНК внутри интерфазных ядер. Действительно, рядом расположенные последовательности должны быть одинаково защищены от мутагенов независимо от их функциональной нагрузки. Такой механизм защиты конкретных генетических локусов от спонтанных и индуцированных мутаций мог бы приводить к мозаичной эволюции генома, предполагаемой в модели Б.Ф.Купа.
Наличие в геноме филогенетически консервативных последовательностей нуклеотидов, как правило, принято объяснять жестким давлением отбора на эти последовательности в процессе эволюции. Например, к числу наиболее консервативных из известных белков относятся гистоны H3 и H4, у которых одна замена на 100 аминокислотных остатков, по-видимому, эволюционно фиксируется в полипептидной цепи не чаще чем в несколько сотен миллионов лет. На противоположном полюсе находятся иммуноглобулины и фибринопептиды, у которых такие мутационные изменения фиксируются в геноме в сотни раз быстрее. В соответствии с традиционной точкой зрения медленно и быстро эволюционирующие белки различаются по содержанию в них функционально значимых сайтов, изменение которых под действием мутаций несовместимо с жизнью. Можно предполагать, однако, что селективная консервативность их генов обеспечивается и структурными особенностями соответствующих локусов генома конкретных видов организмов. Функционирование такого механизма предотвращало бы тяжелые последствия в виде летальных исходов, которыми приходится расплачиваться организмам и соматическим клеткам за мутации в соответствующих локусах, если бы те возникали с такой же частотой, что и в функционально незначимых частях генома. Подобный механизм мог бы быть эффективным в селективной защите от мутаций генетических локусов, содержащих множественные копии генов, так как избыточность генетической информации создает особенно благоприятную почву для дивергенции гомологичных последовательностей. Действительно, дупликации генов и их последующая дивергенция рассматриваются в настоящее время как ключевые механизмы эволюционного процесса.
Еще одна группа данных, свидетельствующих в пользу модели дифференциальной защиты отдельных генетических локусов от спонтанных мутаций, была получена при анализе скоростей образования мутаций в трансгенах, интегрированных в различные участки генома организма-хозяина. Значительную вариабельность в скоростях мутирования локусов мини- и микросателлитов, обнаруженную в геноме млекопитающих и проявляющуюся в различиях уровней их внутрипопуляционного полиморфизма, также частично можно объяснить с помощью обсуждаемой модели.
Таким образом, видоспецифическое пространственное расположение ДНК отдельных генетических локусов интерфазных хромосом создает материальную основу для их дифференциальной чувствительности по отношению к химическим мутагенам экзогенного и эндогенного происхождения. Избирательная доступность индивидуальных нуклеотидов ДНК таких локусов может детерминировать относительную частоту спонтанных мутаций в отдельных локусах геномной ДНК эукариот. Все это может иметь большое значение для онтогенетического и филогенетического развития биологических видов.
|
|