Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Для новых пользователей первый месяц бесплатно. Чат-бот для мастеров и специалистов, который упрощает ведение записей: — Сам записывает клиентов и напоминает им о визите;
— Персонализирует скидки, чаевые, кэшбэк и предоплаты;
— Увеличивает доходимость и помогает больше зарабатывать; Начать пользоваться сервисом

ДНК-гликозилазы и эндонуклеазы клеток микроорганизмов и человека, участвующие в BER
|
|
| Фермент | Источник | Ген | Субстрат (см. рис. I.57) |
| Урацил-ДНК-гликозилаза | E. coli S. cerevisiae Человек | ung UNG UDG | а » » |
| 3-Метиладенин-ДНК-гликозилаза | E. coli » S. cerevisiae Человек | tag alkA MAG MPG | к з, к–м, (б, и) б, к, л к, (д) |
| Fapy/8-оксогуанин-ДНК-гликозилаза (fapy – формамидопиримидин) | E. coli S. cerevisiae Человек | fpg/mutM ? ? | в–д г и/или д » |
| Эндонуклеаза III/тимингликоль-ДНК-гликозилаза | E. coli | nth | в, е, ж |
| Эндонуклеаза VIII | E. coli | nei | » |
| A-G-ДНК-гликозилаза | » Человек | mutY ? | Аденин/в » |
| G-T-ДНК-гликозилаза | » | ? | G-T, (U-G) |
| УФ-эндонуклеаза | T4 M. luteus | ? ? | Пиримидиновые димеры То же |
| Гидроксиметилурацил-ДНК-гликозилаза | Человек | ? | з |
| Формилурацил-ДНК-гликозилаза | » | ? | Ж |
| Примечание. В скобках приведены предположительные субстраты. |

Рис. I.57. Модифицированные азотистые основания ДНК, удаляемые ДНК–гликозилазами при функционировании BER
а – урацил; б – гипоксантин; в – 5–гидроксицитозин; г – 2, 5-диамино-4-формамидопиримидин; д – 7, 8-дигидро-8-оксогуанин; е – мочевина; ж – тимингликоль; з – 5-формилурацил; и – 5-гидроксиметилурацил; к – 3-метиладенин; л – 7-метилгуанин; м – 2-метилцитозин
АР-дезоксирибоза (apurinic/apyrimidinic deoxyribose), образовавшаяся в результате удаления модифицированного азотистого основания апуринового/апиримидинового (AP-) сайта, далее вырезается с помощью АР-лиазы, которая освобождает ее 3’-конец, и АР-эндонуклеазы, гидролизующей ее 5’-концевую фосфодиэфирную связь в АР-сайте (см. рис. I.58). Однонуклеотидная брешь затем заполняется с помощью ДНК-полимеразы, и фосфодиэфирная связь восстанавливается в реакции лигирования. У E. coli репаративный синтез ДНК выполняет ДНК-полимераза I, у дрожжей – ДНК-полимераза d. Из трех ДНК-лигаз, которыми обладают клетки животных, в BER, по-видимому, участвует ДНК-лигаза III.
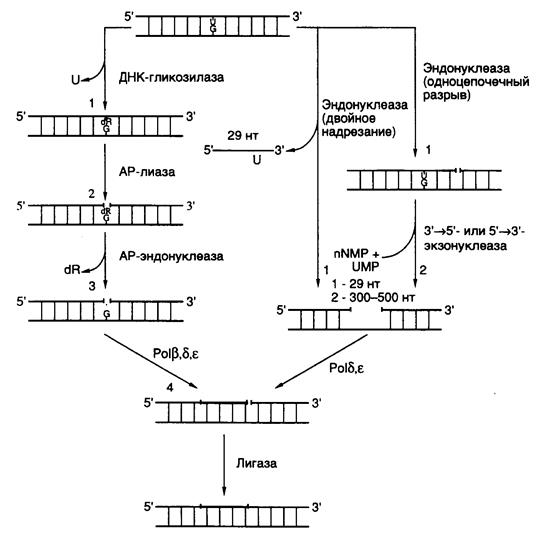
Рис. I.58. Основные пути и этапы эксцизионной репарации у животных
Цифрами обозначены последовательные этапы функционирования BER и NER
В последнее время начаты исследования механизмов сопряжения BER с другими генетическими процессами, протекающими внутри клеток: транскрипцией, репликацией ДНК и регуляцией клеточного цикла. Для соматических клеток менее опасно иметь повреждения ДНК, связанные с появлением некодирующих (AP-) участков, чем ошибочно кодирующих оснований, поскольку последние приводят к образованию мутаций, тогда как первые допускают осуществление полноценной пострепликативной репарации повреждений. ДНК-гликозилазы, участвующие в BER, способны переводить сайты, содержащие модифицированные основания (например урацил), в некодирующие сегменты одной из цепей ДНК. Урацилгликозилазы, ассоциированные с белковыми комплексами репликативных вилок, действуют очень эффективно на одноцепочечные ДНК, и их активность регулируется во время клеточного цикла.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Эксцизионная репарация ДНК путем удаления нуклеотидов (NER). Если в системе BER происходит удаление отдельных поврежденных азотистых оснований ДНК путем разрыва соответствующих N-гликозидных связей между азотистыми основаниями и остатками дезоксирибозы, то в системе NER поврежденные азотистые основания вырезаются в составе олигонуклеотидов. NER может осуществляться двумя путями. В первом случае происходит гидролиз фосфодиэфирной связи по 3’- или 5’-концу на некотором расстоянии от ошибочно спаренного (поврежденного) нуклеотида, который далее целиком удаляется под действием 5’®3’- (или 3’®5’-) экзонуклеазы, гидролизующей цепь ДНК нуклеотид за нуклеотидом в соответствующем направлении от первоначального одноцепочечного разрыва в репарируемой ДНК. Образующаяся брешь далее заполняется ДНК-полимеразой. Такой механизм репарации реализуется у E. coli и человека для вырезания неповрежденных (немодифицированных) ошибочно спаренных нуклеотидов. Механизм последовательного эндо- и экзонуклеазного расщепления ДНК не используется для удаления поврежденных (измененных) нуклеотидов. Это связано, по-видимому, с тем, что такие нуклеотиды (например возникшие в результате образования аддуктов с мутагенами) часто являются ингибиторами экзонуклеаз.
Одним из решений данной проблемы представляется использование ферментной системы, которая вносила бы одноцепочечные разрывы по обе стороны от поврежденного нуклеотида на некотором расстоянии от него с последующим удалением одноцепочечного фрагмента ДНК, содержащего измененный нуклеотид. Действительно, такой второй механизм эксцизионной репарации функционирует у всех исследованных видов живых организмов и будет рассмотрен ниже более подробно.
В универсальном механизме эксцизионной репарации как прокариоты, так и эукариоты гидролизуют 3–5-ю фосфодиэфирную связь с 3'-конца от повреждения (см. рис. I.56). При этом прокариоты гидролизуют также 8-ю связь от 5’-конца измененного нуклеотида, тогда как у эукариотических организмов происходит одноцепочечный разрыв на расстоянии 21–25 нуклеотидов от повреждения со стороны его 5’-конца. Таким образом, прокариоты удаляют измененный нуклеотид в составе 12–13-членных олигомеров, тогда как эукариоты – в составе одноцепочечных фрагментов ДНК длиной в 27–29 нуклеотидов. Ферментная система, вносящая такие двойные одноцепочечные разрывы, получила название эксцизионной нуклеазы (эксцинуклеазы). Образующаяся в молекуле репарируемой ДНК одноцепочечная брешь далее заполняется с помощью ДНК-полимеразы, а фосфодиэфирная связь в остающемся одноцепочечном разрыве восстанавливается ДНК-лигазой.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
Генетика NER. Гены NER E. coli uvrA, uvrB и uvrC не обнаруживают гомологии с соответствующими генами человека. В отличие от них гены NER дрожжей и человека высокогомологичны, и энзимология эксцизионной репарации в этих двух системах также обладает большим сходством. По крайней мере, три заболевания у человека вызываются генетическими нарушениями системы эксцизионной репарации: пигментная ксеродерма, синдром Кокейна и трихотиодистрофия.
Кожа больных пигментной ксеродермой обладает повышенной чувствительностью к дневному свету, что проявляется в виде фотодерматозов, включая рак кожи. В ряде случаев отмечены аномалии нервной системы, причиной которых являются мутации в одном из семи генов: XPA, XPB,...XPG. Однако описаны больные с классическими симптомами пигментной ксеродермы, но с ненарушенной системой NER. Для клеток этих больных характерны изменения в так называемой пострепликативной репарации. Больным с синдромом Кокейна присущи нарушения роста, умственная отсталость, катаракты, повышенная чувствительность к свету с сопутствующими дерматозами. Обнаружены мутации в двух группах генов, приводящие к этому заболеванию. У больных с мутантными генами CS-A или CS-B клетки способны нормально репарировать УФ-повреждения ДНК. У другой группы больных обнаружены мутации в генах XPB, XPD или XPG. У больных трихотиодистрофией со смешанными симптомами выявлены мутации в генах XPB или XPD. Классические симптомы этого заболевания, по-видимому, являются следствием мутации в гене транскрипционного фактора TFIIH.
Получение мутантов с измененной NER у грызунов позволило разбить такие гены на 11 групп комплементации, большинство из которых соответствует группам комплементации XP и CS человека. Часть соответствующих генов человека удалось клонировать, используя их способность исправлять (комплементировать) генетические дефекты в культивируемых мутантных клетках грызунов. Эти гены получили название кросс-комплементирующих генов эксцизионной репарации (ERCC – excision repair cross complementing). Среди них гены XPE и ERCC6 – ERCC11 не требовались для прохождения основных реакций эксцизионной репарации, и их функция неизвестна.
Структура и функции белков NER. В табл. I.21 суммированы некоторые свойства белков животных, участвующих в NER. Большинство таких белков существует in vivo в виде комплексов, поэтому необходимо иметь в виду, что ферментативные активности, обнаруживаемые у отдельных
Таблица I.21
|
|