Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника

Линейные хромосомы бактерий
|
|
Афоризм Жака Моно: " То, что верно для E. coli, – верно и для других бактерий (слона)" получил широкое распространение. К счастью, на деле все обстоит не так скучно. До недавнего времени общепринятым было представление о кольцевой структуре бактериальных хромосом. Однако в 1989 г. была впервые описана у спирохеты Borrelia burgdorfery линейная бактериальная хромосома, которую идентифицировали с помощью электрофореза в импульсном электрическом поле. Размер этого генома составлял всего 960 т.п.о. Вскоре было обнаружено, что линейная и кольцевая хромосомы сосуществуют одновременно у Agrobacteriumtumefaciens, а у грамположительных бактерий рода Streptomyces, обладающих одним из самых больших бактериальных геномов (~8000 т.п.о.), имеется одна линейная хромосома. Представитель актиномицетов Rhodococcusfascians также, по-видимому, обладает линейной хромосомой. Линейные хромосомы у бактерий часто сосуществуют с линейными плазмидами и широко распространены в природе.
Линейные хромосомы и плазмиды наиболее хорошо изученных бактерий рода Streptomyces содержат концевые инвертированные повторы (terminal inverted repeats – TIRs), с которыми ковалентно связаны концевые белки (TP). Несмотря на то что подобные структуры характерны для хромосом аденовирусов и бактериофага f29 Bacillus subtilis, механизм репликации хромосом стрептомицетов существенно отличается от такового вирусных геномов. Если у вирусов синтез ДНК инициируется на конце хромосомы с использованием в качестве затравки TP, ковалентно связанного с нуклеотидом, и продолжается через весь геном до его конца, то репликация хромосомы и линейных плазмид стрептомицетов начинается с внутренней области начала репликации oriC. Синтез ДНК распространяется в обе стороны от области начала репликации по стандартному полуконсервативному механизму и завершается на концах линейных молекул ДНК с образованием 3’-концевых брешей (рис. I.50, а). Наиболее простым решением проблемы заполнения этой бреши могла бы быть прямая инициация репликации теломерных участков хромосом с TP-белка, ковалентно связанного с инициирующим нуклеотидом, что имеет место у аденовирусов (см. рис. I.50, б). Действительно, стрептомицеты используют ТР для репликации теломерных участков, однако механизм распознавания теломер в данном случае существенно отличается. В настоящее время рассматриваются три модели заполнения брешей в теломерных участках линейных хромосом бактерий.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
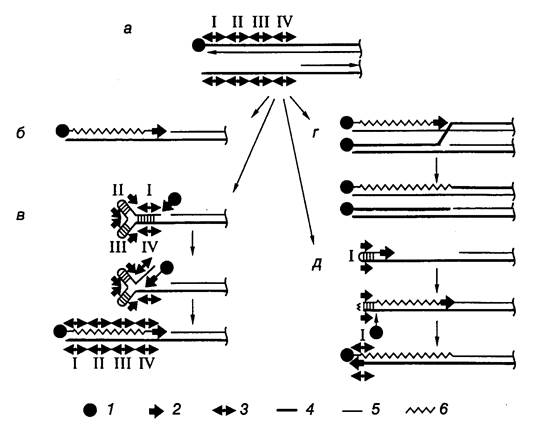
Рис. I.50. Модель достройки теломерных участков хромосом и плазмид Streptomyces
а – структура теломеры после репликации: верхняя цепь ДНК полностью реплицирована, в нижней имеется одноцепочечная брешь, обозначены четыре палиндромные последовательности нуклеотидов; б – маловероятный механизм с участием концевого белка и ДНК-полимеразы; в–д – альтернативные модели репликации, основанные на других механизмах. 1 – концевой белок, 2 – ДНК-полимераза, 3 – палиндром, 4 – родительская цепь ДНК, 5 – дочерняя цепь, 6 – репаративный синтез
В соответствии с первой моделью одноцепочечный участок теломеры, содержащий TIR-последовательность, образует концевую шпильку путем комплементарных взаимодействий нуклеотидов внутренних участков бреши и 3’-концевых нуклеотидов (см. рис. I.50, в). В этом случае синтез ДНК, репарирующий одноцепочечную брешь, инициируется на двухцепочечном участке, образованном палиндромными последовательностями I-IV, с участием ТР и ДНК-полимеразы и продолжается вдоль 3’-концевого одноцепочечного участка хромосомы. Согласно второй модели ТР инициирует репликацию на полностью двухцепочечной дочерней ДНК, вытесняя 5’-концевую цепь родительской ДНК, с которой связан ТР (см. рис. I.50, г). Вытесняемая цепь далее спаривается с выступающим 3’-концом хромосомы, после чего такая разветвленная структура разрешается с помощью гомологичной рекомбинации. Эта модель предполагает участие в заполнении брешей белка RecA (для переноса цепи ДНК) и продуктов генов ruv (для разрешения структуры Холидея), что подтверждается генетическими данными. В третьей модели одноцепочечный палиндром I образует шпильку, 3’-конец которой служит затравкой для синтеза ДНК, в результате которого заполняется брешь (см. рис. I.50, д). ТР образует одноцепочечный разрыв напротив первоначального 3’-конца, который является затравкой для последующего синтеза ДНК. В результате шпилька разворачивается и восстанавливается структура теломеры. Эта модель аналогична модели " катящейся шпильки", предложенной для объяснения механизма репликации генома парвовирусов. В данной модели роль ТР отличается от его функций в качестве белка-затравки в рассмотренных выше примерах.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
Неизвестно, как много форм линейных бактериальных хромосом существует в природе. Не изучены и таксономические проблемы, связанные с топологией хромосом в царстве эубактерий. Если каждый тип хромосом характерен для отдельного таксономического домена, то можно предполагать, что топология хромосом играет важную роль в эволюции бактерий. Альтернативно топологические взаимопревращения хромосом могут быть относительно частыми событиями, а линейные и кольцевые хромосомы присутствуют только у близких видов бактерий. Нестабильность хромосом стрептомицетов (образование протяженных делеций и амплификация последовательностей нуклеотидов) недавно стали связывать с перестройками в их концевых участках, часть из которых сопровождалась образованием кольцевых хромосом. Таким образом, эволюционная роль топологии бактериальных хромосом может быть определена только в результате будущих исследований.
|
|