Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника

Нуклеоид бактериальной клетки
|
|
Каждый, кому приходилось разрушать бактериальные клетки в мягких условиях, например с помощью лизоцима или детергентов, наблюдал
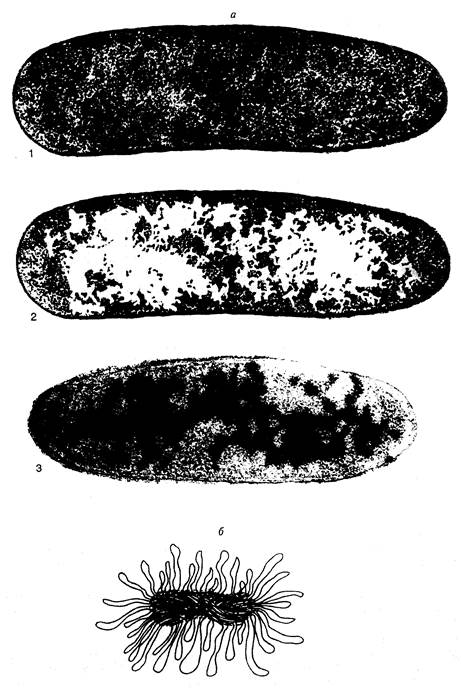
Рис. I.1. Нуклеоид E. coli
а – электронно-микроскопические фотографии срезов бактериальных клеток, полученные методом криофиксации. 1 и 2 – один и тот же снимок (последний ретуширован). На фотографии 2 белыми пятнами отмечены области цитоплазмы, свободные от рибосом. На фотографии 3 видны молекулы ДНК, специфически окрашенные с помощью антител;
б – модель нуклеоида в функционально-активном состоянии А. Райтера и А. Чанга. Изображены многочисленные петли активно транскрибируемой ДНК
замечательную картину превращения легко подвижной суспензии бактериальных клеток в вязкую желеобразную массу, простое перемешивание которой требует усилий. Это происходит из-за того, что компактно упакованные гигантские хромосомы бактериальных клеток (длина хромосомной ДНК E. coli составляет ~4, 6 млн. п.о.) после разрушения оболочки клеток выходят в окружающую среду и свободно в ней распределяются. В лизатах бактериальных клеток их ДНК прочно ассоциированы с белками, освобождение от которых требует проведения многократных фенольных депротеинизаций. Такой простой опыт наглядно указывает на то, что в бактериальных клетках их единственная хромосома сильно компактизована и, возможно, пространственно упорядочена.
Электронно-микроскопическое изучение срезов бактериальных клеток в разных условиях и более ранние исследования бактерий с помощью светового микроскопа продемонстрировали компактное распределение ДНК в бактериальной клетке. Поскольку такие структуры отдаленно напоминали ядра эукариот, они получили название нуклеоидов, или ДНК-плазмы. Эти термины подчеркивают генетические функции нуклеоида, но в то же время и существенные морфологические отличия от обычных интерфазных ядер эукариот, прежде всего, отсутствие ядерной оболочки, которая бы отделяла гены бактерии от окружающей их цитоплазмы. Исследование бактериальных клеток с помощью электронной микроскопии в мягких условиях без предварительной химической фиксации показало, что нуклеоиды представлены в виде диффузно окрашенных областей, свободных от рибосом (рис. I.1, а). При этом вытянутые участки ДНК на внешней части нуклеоидов направлены в окружающую цитоплазму. С помощью специфических антител установлено, что молекулы РНК-полимеразы, ДНК-топоизомеразы I и гистоноподобного белка HU ассоциированы с нуклеоидами. Вытянутые участки ДНК по периферии нуклеоидов обычно интерпретируют как сегменты бактериальной хромосомы, вовлеченные в транскрипцию. Полагают, что эти участки состоят из петель ДНК бактериальной хромосомы, которые в зависимости от физиологического состояния клетки находятся в транскрипционно-активном состоянии или втягиваются внутрь нуклеоидов при подавлении транскрипции. Модель функционально-активного нуклеоида А.Райтера и А.Чанга представлена на рис. I.1, б. По мнению авторов, размытая структура поверхности нуклеоидов, видимая под электронным микроскопом, отражает подвижное состояние активно транскрибируемых петель ДНК. В этой модели четко прослеживается аналогия со структурой хромосом типа ламповых щеток у животных.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Таким образом, нуклеоид бактериальных клеток не является статическим внутриклеточным образованием или компартментом, которые можно четко определять морфологически. Напротив, во время различных фаз роста бактериальных клеток нуклеоид непрерывно меняет форму, что, по-видимому, сопряжено с транскрипционной активностью определенных бактериальных генов. Так же как и в хромосомах эукариот, ДНК нуклеоида ассоциирована со многими ДНК-связывающими белками, в частности гистоноподобными белками HU, H-NS и IHF, а также топоизомеразами, которые оказывают большое влияние на функционирование бактериальных хромосом и их внутриклеточную компактизацию. Однако детальные молекулярные механизмы конденсации бактериальной ДНК с образованием лабильных " компактосом " (по аналогии со стабильными нуклеосомами эукариот) пока неизвестны. В последнее время возрастает интерес к бактериальному так называемому LP-хроматину (low protein chromatin), для которого характерно относительно низкое содержание белкового компонента. Аналогичный LP-хроматин обнаруживают у вирусов, в митохондриях, пластидах и у динофлагеллят (жгутиконосцев). Следовательно, этот тип структурной организации генетического материала претендует на универсальность и ассоциирован с определенными формами регуляции экспрессии генов, свойственными прокариотическим организмам.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
В последние годы наблюдается прогресс в исследовании первичной структуры бактериальных хромосом. Определена полная последовательность нуклеотидов хромосом паразитических бактерий: микоплазмы Mycoplasma genitalium и Haemophilus influenzae. В 1997 г. усилиями интернационального коллектива ученых была определена полная первичная структура хромосом E. coli и Bacillus subtilis длиной в ~4, 6 и 4, 2 млн п.о. соответственно Все это позволяет надеяться, что в ближайшее время произойдут новые открытия в области исследований структуры бактериальных геномов и функционирования их генов.
|
|