Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника

Репликация кольцевых ДНК с образованием «глазка». q-структура
|
|
В случае репликации кольцевых молекул ДНК присутствие глазка приводит к образованию q (тэта)-структуры, изображенной на рис 4.2.
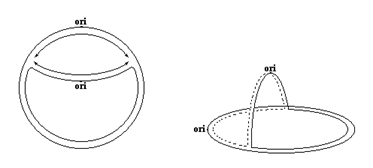
| Рис. 4.2 | В кольцевых молекулах ДНК репликационный глазок образует q-структуру. Приведены две проекции реплицирующейся кольцевой ДНК. Символом ori обозначена точка начала репликации, стрелками показано направление репликации. |
Последовательные стадии репликации кольцевой ДНК вируса полиомы, зафиксированные под электронным микроскопом, приведены на рис. 4.3. На отдельных микрофотографиях хорошо видно, что появившийся глазок постепенно расширяется, формируя q-структуру. Нереплицированный сегмент уменьшается, что завершается разделением двух дочерних молекул ДНК.
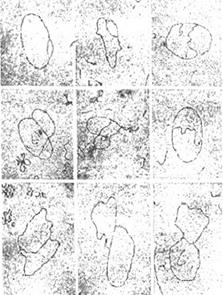
| Рис. 4.3 | Репликационный глазок увеличивается по мере продвижения репликативных вилок вдоль репликона (Lewin B., Genes III, New York, Wiley, 1987).
Репликация по типу катящегося кольца
До сих пор мы рассматривали возможность репликации в одном или двух направлениях, когда одновременно синтезируются обе новые цепи ДНК. Как в линейной, так и в кольцевой молекуле ДНК (например, у E. coli) движение репликативной вилки (или вилок) формирует глазок. Если матрицей служит кольцевая ДНК, реплицирующаяся молекула принимает форму q-структуры.
Однако возможен и другой вариант репликации кольцевой молекулы ДНК. В этом случае двухцепочечная кольцевая ДНК надрезается специфическим ферментом в уникальном сайте только одной цепи, и к образовавшемуся в результате надреза 3`-OH-концу с участием ДНК-полимеразы III присоединяются дезоксинуклеотиды. Вследствие того, что матрицей служит интактная замкнутая цепь ДНК, в вилке синтезируется только лидирующая цепь [8]. По мере синтеза лидирующей цепи происходит вытеснение 5/-конца исходной надрезанной родительской цепи (рис. 4.10).
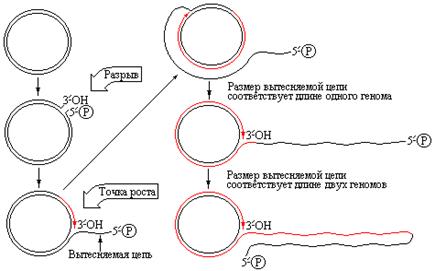
Такой тип структуры получил название катящегося кольца, поскольку точка роста скользит вокруг кольцевой матричной цепи. Так как вновь синтезируемый материал ковалентно связан с исходным материалом, вытесняемая нить достигает больших размеров и содержит любое число единиц геномов, синтезируемых путем непрерывного вращения матрицы. Катящееся кольцо используется in vivo многократно. Для чего это нужно? У фагов М13 или jХ174, чьи зрелые геномы представлены одноцепочечными кольцевыми ДНК, указанный способ репликации используется на поздних стадиях инфекционного процесса, после того как геномная кольцевая ДНК превращается в двухцепочечную репликативную форму. Постоянно отделяющиеся в результате репликации одиночные цепи ДНК приводят к образованию серии геномов (отдельные цепи могут включать от двух до пяти фаговых геномов), которые затем фрагментируются в каждой точке начала репликации и встраиваются в фаговые частицы в виде одноцепочечных замкнутых зрелых форм. Кроме того, возможно их использование в дальнейших циклах репликации. Иногда освобождаемая отдельная цепь катящегося кольца может быть превращена в двухцепочечную ДНК путем синтеза комплементарной цепи, при этом образуются двухцепочечные конкатемерные молекулы, необходимые для созревания определенных фаговых ДНК, например, ДНК фага l. Фаг l использует такой способ репликации при образовании своей линейной двухцепочечной геномной ДНК. На ранних стадиях инфицирования линейная вирусная ДНК превращается в кольцевую репликативную форму. Для образования линейной ДНК фагового потомства двухцепочечные кольца надрезаются и асимметрично реплицируются, как это происходит в случае ДНК фагов М13 и jХ174. Однако вытесняемые одиночные цепи превращаются в двухцепочечные структуры (рис. 4.11). Забиваем Сайты В ТОП КУВАЛДОЙ - Уникальные возможности от SeoHammer
Каждая ссылка анализируется по трем пакетам оценки: SEO, Трафик и SMM.
SeoHammer делает продвижение сайта прозрачным и простым занятием.
Ссылки, вечные ссылки, статьи, упоминания, пресс-релизы - используйте по максимуму потенциал SeoHammer для продвижения вашего сайта.
Что умеет делать SeoHammer
— Продвижение в один клик, интеллектуальный подбор запросов, покупка самых лучших ссылок с высокой степенью качества у лучших бирж ссылок. — Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта. — Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы). — SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание. SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
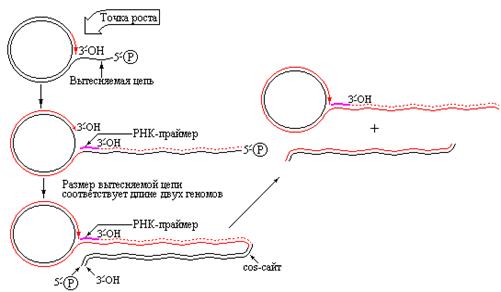
Сначала во множестве сайтов вдоль одиночной цепи праймаза синтезирует короткие фрагменты РНК, которые используются ДНК-полимеразой III в качестве праймеров. Затем РНК удаляется с помощью ДНК-полимеразы I и бреши лигируются с участием ДНК-лигазы. При упаковке ДНК в фаговые частицы в специальных участках конкатемеров, называемых cos-сайтами и отстоящих друг от друга на длину вирусного генома, происходит разрезание. В результате длинные дуплексы многократно повторенной ДНК фага l расчленяются на фрагменты, соответствующие по размеру зрелой ДНК, обнаруживаемой в вирионах [9]. Следует отметить, что данный способ образования конкатемерных молекул ДНК не единственный. Такие молекулы образуются при размножении фага Т4. Механизм катящегося кольца используется при образовании большого количества копий кольцевых молекул рДНК (кодирующих рРНК) в ооцитах Xenopus laevis.
|
|
|