Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Распознавание грамотрицательных бактерий
|
|
Схема клеточной стенки грамотрицательной бактерии и основных структур – мишеней системы врожденного иммунитета - представлена на рисунке 9. Многие важные для микробной клетки вещества уже рассмотрены выше (пептидогликан, бактериальная ДНК). Важнейшим отличительным элементом грамотрицательной бактерии является липополисахарид (ЛПС).

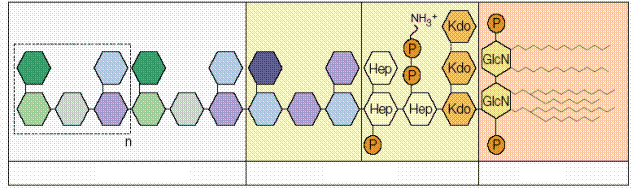
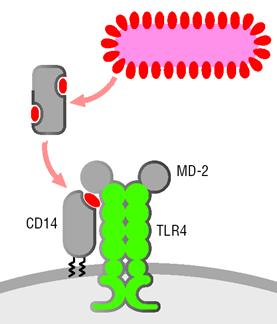
ЛПС состоит из гидрофобной (липид А) и гидрофильной (центральный полисахарид и О-антиген) частей, является основным структурным элементом внешней псевдомембраны грамотрицательных микробов и проявляет выраженную биологическую активность. Иммунная система распознает ЛПС с помощью множества различных рецепторов: CD14, интегрины (CD11c/CD18, CD11b/CD18), рецепторы комплемента CR3 и CR4, CD55, белки теплового шока hsp70 и hsp90, рецепторы к хемокинам (CXCR4), TREM-1, RP105, фактор активации тромбоцитов ADAM10. К наиболее важным относят рецепторный комплекс ЛПССБ-CD14-ТПР4. ЛПС-СБ (липополисахарид связывающий белок) является белком острой фазы воспаления, синтезируется гепатоцитами в ответ на ИЛ6 и цитокины воспаления, связывается с ЛПС и выполняет опсонизирующую функцию. ЛПССБ контролирует содержание ЛПС в крови, способствует фагоцитозу бактерий и доставляет ЛПС к сигнальному рецептору ТПР4. CD14 - это мембранный гликопротеин, экспрессирующийся на моноцитах и макрофагах. CD14 существует и в растворимой форме. рCD14 способен связывать ЛПС и активировать клетки эндотелия/эпителия.
 В зависимости от структуры (особенно, липида А) и распознающих рецепторов, а также концентрации, ЛПС способен индуцировать мощный воспалительный ответ, выделение больших количеств ФНО-альфа, ИЛ1; ИЛ12, ИФН-гамма, что (в условиях эксперимента или при сепсисе) может привести к шоку и полиорганной недостаточности, умеренный воспалительный ответ, или не возбуждает активности иммунной системы. Сильное воспаление приводит к усилению защитных и повреждающих реакций, а также к развитию адаптивного иммунного ответа (подобный эффект часто использовался для приготовления иммуномодуляторов и адьювантов).
В зависимости от структуры (особенно, липида А) и распознающих рецепторов, а также концентрации, ЛПС способен индуцировать мощный воспалительный ответ, выделение больших количеств ФНО-альфа, ИЛ1; ИЛ12, ИФН-гамма, что (в условиях эксперимента или при сепсисе) может привести к шоку и полиорганной недостаточности, умеренный воспалительный ответ, или не возбуждает активности иммунной системы. Сильное воспаление приводит к усилению защитных и повреждающих реакций, а также к развитию адаптивного иммунного ответа (подобный эффект часто использовался для приготовления иммуномодуляторов и адьювантов).
Флагеллин. Флагеллин является основной составляющей бактериальных жгутиков и важным объектом внимания врожденного иммунитета. За распознавание флагеллина отвечает ТПР5, который экспрессирован на клетках эпителия, моноцитах, незрелых дедритных клетках. При этом ТПР5 на эпителиоцитах экспрессируется полярно, на базо-латеральной поверхности, что вызывает его активацию только при проникновении бактерий через эпителиальный барьер. Стимуляция ТПР5 на клетках эпителия приводит к их активации, повышению экспрессии молекул адгезии, секреции хемокинов, специфичных для нейтрофилов и дендритных клеток, продукции антибактериальных веществ и генерации микроокружения, способствующего развитию Th2 клеток. Активация ТПР5 на дендритных клетках приводит к стимуляции гуморального иммунного ответа (в частности, продукции IgA).
Фимбрии Фимбрии различных бактерий (E.coli, P.gingivalis, Campylobacter jejuni, Shigella flexneri) играют важную роль в адгезии и колонизации эпителия, а также распознаются рецепторами системы врожденного иммунитета, в частности, ТПР2 и 4. Распознавание фимбрий играет важную роль в организации врожденного иммунитета слизистых оболочек.
Энтеротоксины возбудителей инфекций ЖКТ (например, LT-II Е.coli) также могут распознаваться через ТПР (ТПР2).
Необходимо учитывать, что в реальной ситуации в клетке генерируется большое количество сигналов о распознавании, к примеру, ЛПС, пептидогликана, ЛТК, флагеллина, бактериальных нуклеиновых кислот и т.д., которые синнергично усиливают воспалительную составляющую ответа и обусловливают его индивидуальность.
Нормальная микрофлора. Несмотря на наличие доказанных различий в структурных компонентах патогенных и непатогенных бактерий, они высококонсервативны и, как правило, экспрессируются широким кругом микроорганизмов. Таким образом, неизбежно должны существовать механизмы, ограничивающие распознавание/реактивность в ответ на микроорганизмы нормальной микрофлоры: снижение апикальной экспрессии ТПР в обычных условиях; нефункциональность рецепторов; смещение экспрессии ТПР на базальную (латеральную) поверхность, поверхность крипт, внутриклеточные везикулы (пузыри комплекса Гольджи); смещение максимума экспрессии на клетки субэпителиальных тканей; повышение реактивности отдельно взятых рецепторных комплексов (активация в ответ на флагеллин или материал фимбрий). Отдельно необходимо отметить существование негативной регуляции сигнальных рецепторов (ТПР): определенную роль играют растворимые формы ТПР (регулируют активность лигандов и рецепторных комплексов), однако более важны внутриклеточные механизмы (белки TOLLIP, IRAKm, TRIAD3A и др).
|
|