Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Психомоторной деятельности
|
|
Собственно двигательная зона коры, известная, как было уже отмечено, со времени ее открытия G. Frisch u. E. Hitzig (1870), расположена спереди от центральной борозды, в так называемой прецентральной области передних отделов коры. Эта область, как установлено, дифференцировалась в ходе филогенеза из единой сензомоторной коры и распалась на два поля: первичное, или моторное (поле 4), представляющее собой центральное двигательное поле, и вторичные, или премоторные (поля 6 и 8), являющиеся периферическими двигательными полями (рис. 2). Передняя центральная извилина, включающая в себя поля 4 и 6, является корковой областью двигательных проекций.
Многочисленными экспериментальными исследованиями выявлено, что раздражение коры передней центральной извилины в какой-либо точке вызывает сокращение соответствующих мышц, то есть их движение.
Литературные данные свидетельствуют, что общая структурно-функциональная характеристика названных выше полей и их связи с подкорковыми образованиями мозга в целом сходны с первичными и вторичными полями задних отделов мозга, в частности кожно-кинестетического анализатора (поля 3, 1, 2), рассмотренного нами в предыдущем разделе этой главы. Так, характер протяженности и объем двигательной зоны соматотопической проекции частей и органов тела в первичном поле 4 сходен с аналогичными характеристиками центрального поля 3 кожно-кинестетической (сенсорной) ядерной зоны. А именно установлено, что в зависимости от функциональной нагрузки, от того, насколько тонкие и дифференцированные движения осуществляет та или другая мышца, ее проекция будет занимать больше или меньше места. Например, выявлено, что мелкие мышцы каждой фаланги всех пальцев руки имеют отдельное представительство в передней центральной извилине, в то время как крупные мышечные группы ног и туловища представлены суммарно (рис. 1 и 3).
Вместе с тем, как показывают специальные исследования, в работе передних и задних отделов мозга выявлены и существенные различия, заключающиеся в том, что " в организации коры всей лобной доли мозга, в отличие от задней части полушария, основной акцент падает на осуществление координированных и целенаправленных воздействий организма на внешний мир в ответ на воспринимаемые комплексы раздражений" (А.Р. Лурия, 1962, С.48).
В строении первичного поля 4 взрослого человека отмечаются, как установлено, две структурные особенности: во-первых, наличие мощно развитых гигантских клеток Беца в слое V, дающих начало наиболее крупным волокнам пирамидного пути, которые проводят импульсы произвольных движений на двигательные центры скелетной мускулатуры, расположенные в стволе головного мозга и спинном мозге; во-вторых, отсутствие отчетливо обособленного, как в задних отделах полушарий слоя IV, приспособленного для восприятия афферентных импульсов из подкорки, а наличие для выполнения этих функций лишь вкрапленных в гигантопирамидное поле 4 рассеянно расположенных соответствующих нервных клеток.
Обозначенные структурные особенности " свидетельствуют о том, что в центральном двигательном поле сравнительно наибольшего развития достигают элементы нейронного комплекса, приспособленного для максимально быстрого и кратчайшим путем проведения произвольных импульсов на эффекторные нейроны центральной нервной системы" (А.Р. Лурия, 1962; С.48).
Указанные особенности микроскопической структуры поля 4 обусловливают, по мнению исследователей, его физиологическую и клиническую характеристику, сводящуюся, как было выявлено, к весьма дробно дифференцированному у человека представительству в этом поле движений отдельных мышечных групп.
Вторичное премоторное поле 6, как свидетельствуют данные физиологических и клинических исследований, отвечают за выполнение и автоматизацию более сложных координированных движений, двигательных комплексов, протекающих во времени и вовлекающих в совместную деятельность различные группы мышц тела.
По сравнению с гигантопирамидным полем 4 цитоархитектонически поле 6 отличается отсутствием гигантских клеток Беца и лучше развитыми пирамидами слоя III. Проекционные связи премоторной коры с подкорковыми образованиями составляют, как известно, важную часть экстрапирамидных систем коры, которые " в отличие от прямого пирамидного пути достигают конечных двигательных центров головного и спинного мозга через ряд переключений в лежащих ниже коры уровнях центральной нервной системы" (А.Р. Лурия, 1962; С.49). Пирамидная и экстрапирамидная системы наглядно представлены схемой J.W. Papez, 1958 (рис. 7).
К периферическому (вторичному) отделу двигательной коры относят также поле 8, являющееся глазодвигательным полем. В цитоархитектоническом строении этого поля обнаруживаются признаки переходности к полям собственно лобной коры. Поле 8 отвечает за осуществление координированных движений взора, его фиксации при манипулировании предметами под контролем зрения.
Моторное 4 и премоторные 6 и 8 поля прецентральной области, составляющие, как известно, единый кинетический комплекс коры, имеют хорошо развитую проекционную и ассоциационную афферентацию в виде двусторонних связей между собой, с другими полями коры и с подкорковыми образованиями. Однако афферентация, направляющаяся в моторную и премоторную кору, имеет иные источники и переключается в иной группе ядер зрительного бугра, чем афферентация, поступающая в корковые отделы задней части полушария.
Основными каналами импульсов, идущих в поля прецентральной области, являются пути от мозжечка через красное ядро и зрительный бугор на кору (рис. 7). Доказано, что эта афферентация служит для обратного замыкания импульсов, которые циркулируют по экстрапирамидным корково-подкорковым системам мозга
Выявлено, что специфика проекционной афферентации двигательной коры обусловлена ее функциональной ролью в совокупной деятельности корковых концов анализаторов. Принцип обратной связи, лежащий в основе рефлекторной деятельности центральной нервной системы, реализуется, как известно, в ядерных зонах анализаторов с их зонами перекрытия в процессе непосредственного восприятия анализаторами различных по сложности раздражений, поступающих извне Установка и настройка анализаторов на адекватное взаимодействие человека с действительностью обеспечивается как раз двигательным анализатором, выполняющим интегративную функцию.
Применительно к работе двигательной коры, ответственной, как известно, за организацию, " программирование" и реализацию произвольной двигательной активности человека, обратная связь выступает в качестве основного источника информации об эффекте совершаемых движений и действии Роль двигательной коры, таким образом, своди 1ся в общих чертах к сопоставлению " заданной программы" двигательною акта, формирующейся в коре задних отделов мозга, с реальным ее выполнением, то есть к определению соответствия между программой и выполняемым действием и внесению необходимых и своевременных коррекции по ходу реализации этих действий (НА Бернштейн, 1947, К.Н Pubram, 1959, 1960 и др.) Вполне очевидно, что расстройство согласованной работы этой системы, связанное с различными по локализации поражениями или незрелостью мозговых cтруктyp, будет вызывать в каждом конкретном случае специфические двигательные нарушения в различных группах мышц, влияя 1ем самым на успешность организации и осуществления соответствующей психомоторной деятельности человека.
Из сказанного становится понятным значение нормального функционирования эфферентных (центробежных) и афферентных (центростремительных, осуществляющих обратную связь) цепей переключений импульсов, составляющих зкстрапирамидную систему мозга и соединяющих двигательную зону с подкорковыми образованиями (рис 7), в осуществлении каждою произвольного двигательного акта, в координированном протекании психомоторной деятельности человека в целом
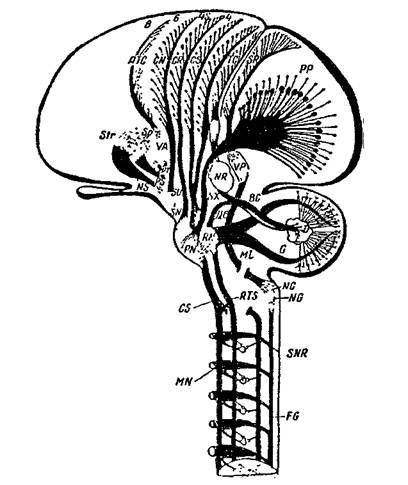
Рис.7. Схема пирамидной и экстрапирамидной систем (по J.W. Papez; 1958)
FG-NG-ML- VP-SR - проводящие пути и переключения кожнокинестетических
рецепции, FF-PP} D-BC-NR-VL-TC- связи коры мозга с мозжечком,
CS-MN - корково-двигательный пирамидный путь, CS-RN-RTS - связи коры с ядрами ретикулярной формации,
CR-SN- связи коры с черным веществом, Sir - подкорковые узлы больших полушарий
Установлено, что подобно тому, как в процессе филогенеза усложнение функциональных отношений между анализаторами задних отделов коры вызывает взаимное структурное перекрытие ядерных зон с образованием особых " зон перекрытия" корковых концов анализаторов (третичных полей коры), дальнейшее развитие и совершенствование всей сферы произвольных, направленных во внешний мир двигательных актов человека обусловило формирование третичных полей собственно лобной области. Эти поля, как было установлено, занимая у человека около четверти всей протяженности лобной коры, сформировались в результате разрастания кпереди (к лобному полюсу) элементов того типа нейронной организации, который заложен в основе двигательной зоны коры. Однако, в отличие от полей двигательной юны в полях лобной области обнаружено более отчетливое обособление слоя IV, что, по замечанию Г.И. Полякова, " может указывать на более компактный характер афферентной сигнализации, поступающей в лобную кору от соответствующей группы ядер зрительного бугра" (А.Р. Лурия, 1962; С.51).
Третичные поля лобной области, расположенные кпереди от моторной (поле 4) и премоторной (поля 6 и 8) зон, составляют, так называемые префронтальные отделы коры, которые, как известно, включают в свой состав ряд образований (поля 9, 10, 11, 45, 46). Часть этих полей находится на конвекситальной, а часть на медиобазальной поверхности лобных долей (рис. 2 и 6).
По мнению А.Р. Лурия (1962, 1969), генетическая связь лобной коры с двигательной корой полей 4 и 6 " подтверждает тот важный факт., что кора лобной области вместе с моторной и премоторной зоной может быть с полным основанием отнесена к корковым отделам двигательного анализатора" (1962; С. 196; выделено А.Л.).
Особенности строения, отличающие префронтальные отделы коры от полей 4 и 6, обусловливают, как указывает А.Р. Лурия, своеобразие функций лобных долей. Особенности их цитоархитектоники, как известно, состоят в том, что в префронтальных отделах отсутствуют гигантские пирамидные клетки Беца и более мощно развиты второй и третий (ассоциативные) слои, нейроны которых обладают более тонким строением. Также отмечается существенное отличие системы вертикальных связей этих отделов с нижележащими отделами зрительного 6yгpa от связей " коркового ядра" двигательного анализатора.
Сложность нейронного строения полей префронтальной области подтверждается нейронографическими исследованиями многих отечественных и зарубежных авторов (Е.П. Кононова, 1940, 1948; Г.И. Поляков, 1959; И.Н. Филимонов, 1957 и др.; W.S. Mc'Culloch, 1943; О. Sugar, J.D. French, J.G. Ghusid, 1948 и др.). Кроме того, исследованиями Московского института мозга установлено, что если на первых стадиях внеутробного онтогенеза поля " коркового ядра" двигательного анализатора (4, 6, 8,) существенно опережают по темпу развития поля префронтальнои области, то на последующих стадиях онтогенеза поля префронтальнои области начинают развиваться значительно быстрее полей заднелобных отделов, и территория, занимаемая ими, стремительно возрастает.
Выявлено также, что проекционные центростремительные и центробежные связи лобной коры устанавливаются со многими подкорковыми образованиями, но особенно значительного развития достигают у человека связи с мозжечковой системой, что обусловлено, по мнению исследователей, прямохождением и значением мозжечковых координации для осуществления целенаправленных действий. Вместе с тем установлено, что поля лобной области обладают обширными двусторонними ассоциационнъши связями с полями как прецентральной области, так и других областей коры, расположенных позади центральной борозды, а также на внутренней и нижней поверхностях полушарий (А.Р. Лурия, 1962, 1969; Г.И.Поляков, 1959 и др.; J.C. Dusser de Barrenes a. W.S. Mc'Culloch, 1941; W.S. Mc'Culloch, 1943; O. Sugar, J.D. Frensch, J.G. Ghusid, 1948 и др.). Доказано, например, что глазодвигательное поле 8 обладает специфическими афферентно-эфферентными связями с полями зрительных отделов, а поля 12, 45, 46 имеют аналогичные афферентно-эфферентные связи с полями 22, 37, 39 теменно-височной области, которые входят в систему речевых зон. Интересные сводные данные нейронографических исследований связей между полями префронтальнои коры и другими корковыми полями указанных выше авторов представлены А.Р. Лурия в табличном изложении (1962; С. 197; Табл. 1). Эти сведения, раскрывая афферентные и эфферентные связи между различными корковыми полями, дают общее представление о множественных ассоциационных связях различных отделов лобной коры не только между собой, но и с разными отделами зацентральных областей мозговой коры, что позволяет, на наш взгляд, при анализе тех или иных отклонений в моторной сфере видеть возможные взаимовлияния и взаимосвязи этих отклонений с другими психическими функциями и целыми функциональными системами, а также представлять возможную структуру нарушений и механизмы их возникновения.
Анализ обширных литературных данных о сложном строении префронтальных отделов коры головного мозга, о их многообразных связях с другими отделами больших полушарий, а также результаты собственных исследований позволили А.Р. Лурия (1962, 1969) не только сделать заключение о принадлежности префронтальных областей к системе корковых отделов двигательного анализатора, но и подвели к предположительным выводам о их важной роли в афферентации движений. " Принимая афферентные импульсы едва ли не от всех важных разделов коры головного мозга, они, по-видимому, играют существенную роль в переработке этих импульсов и передаче их на систему двигательного анализатора" (А.Р. Лурия, 1962; С.197).
Полученные фактические данные о том, что кора лобной области по своему строению близка к моторным и премоторным областям и практически входит в систему центральных отделов двигательного анализатора позволили в свое время А.Р. Лурия высказать некоторые предположения о роли лобных долей мозга в организации психической деятельности человека, о ее ближайшем участии " в формировании анализа и синтеза тех возбуждений, которые лежат в основе двигательных процессов" (1962; С.99).
Им высказана мысль о том, что " лобные доли мозга объединяют информацию о внешнем мире, поступающую через аппараты экстерорецепторов, и информацию о внутренних состояниях организма, и являются аппаратом, позволяющим регулировать поведение организма на основе учета эффекта совершаемых им действий" (гам же, выделено А.Л.).
|
|