Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Для новых пользователей первый месяц бесплатно. Чат-бот для мастеров и специалистов, который упрощает ведение записей: — Сам записывает клиентов и напоминает им о визите;
— Персонализирует скидки, чаевые, кэшбэк и предоплаты;
— Увеличивает доходимость и помогает больше зарабатывать; Начать пользоваться сервисом

Біохімія м’язів і м’язового скорочення
|
|
Лекція № 1
ПЛАН
1. Типи м’язових волокон.
2. Структурна організація м’язових волокон.
3. Хімічний склад м’язових тканин.
4. Структурні і біохімічні зміни в м’язах при скороченні і розслабленні.
5. Молекулярний механізм м’язового скорочення.
М’язи є молекулярною системою, в якій відбувається трансформація хімічної енергії АТФ у механічну енергію скорочення та руху.
На м’язи припадає 40-45% маси тіла. За своїми властивостями м’язи характеризуються великою еластичністю, пластичністю та скоротливістю. Це єдина унікальна природна система, наділена здатністю перетворювати безпосередньо хімічну енергію в механічну з високим коефіцієнтом корисної дії.
Морфологічно м’язи у хребетних тварин поділяють на поперечносмугасті, або скелетні, гладенькі та серцевий м’яз (міокард). Перші під мікроскопом мають вигляд довгих волокон, в яких регулярно чергуються світлі й темні смуги. Другі складаються з коротких волокон, що не містять смуг (рис. 1) міокард є перехідною формою. Вони відрізняються морфологічними, біохімічними і функціональними особливостями, а також шляхами розвитку.
Скорочення гладеньких м’язів ініціюється нервовими імпульсами, певними гормонами і не залежить від волі людини, так як їїх тонус не контролюється нашою свідомістю.
Серцевий м’яз скорочується ритмічно з послідовно змінюючимися циклами скорочення (систола) і розслаблення (діастола) незалежно від волі людини його сокрочення регулюється гормонами, наприклад катехоламінами.
Скелетні м’язи прикріплені в основному до кісток, що зумовило їх назву. Скорочення скелетних м’язів ініціюється нервовими імпульсами і керується свідомістю людини.
Відповідно до певних ультраструктурних і метаболічних особливостей, скелетні м’язи поділяють на два типи:
- червоні (повільні) скелетні м’язи;
- білі (швидкі) скелетні м’язи.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Червоні м’язи — тип м’язів, що добре забезпечуються кров’ю та містять багато міоглобіну — (О2-зв’язуючого білка який утворює резерви кисню в м’язових клітинах. В червоних м’язах багато мітохондрій, і вони мають високу здатність до окислювальних процесів, використовуючи як субстрат глюкозу, жирні кислоти, кетонові тіла. Швидкість скорочення складає більш ніж 110 мс. Цей тип м’язів найбільш пристосований до довготривалої фізичної роботи, але включення енергетичних резервів у них відбувається повільно.
Білі м’язи — тип м’язів, що містять невелику кількість мітохондрій; завдяки високій активності гліколітичних ферментів (глікогенфосфорилаз) вони в більшій мірі пристосовані до отримання енергії при анаеробному розщепленні глікогену. За рахунок анаеробного глікогенолізу та гліколізу білі м’язи більш швидко (50 мс), ніж червоні, переходять до максимальної активності з високою частотою скорочень, але швидше втомлюються.
М’язова тканина тварин і людини містить від 73 до 78 % води. Приблизно 22-27 % від маси м’яза припадає на частку сухого залишку, переважно білків. Крім білків, у м’язах знаходяться глікоген та інші вуглеводи, різні ліпіди, екстрактивні речовини та мінеральні солі.
Таблиця 1. Хімічний склад поперечносмугастих м’язів
| Речовина | Вміст (% на сиру масу) |
| Вода | 73-78 |
| Сухий залишок | 22-27 |
| У тому числі: | |
| Білки | 17-21 |
| Глікоген | 0, 5-3, 0 |
| Фосфоліпіди | 0, 02-1, 0 |
| Холестерин | 0, 02-0, 23 |
| Креатинін | 0, 003-0, 005 |
| АТФ | 0, 25-0, 40 |
| Креатин+креатинфосфат | 0, 2-0, 55 |
| Карпозин | 0, 2-0, 3 |
| Молочна кислота | 0, 01-0, 02 |
| Неорганічні речовини | 1, 0-1, 5 |
В м’язах розрізняють 3 види білків: білки саркоплазми, білки міофібрил і білки строми.
У саркоплазмі м’язів містяться білки, що розчиняються у воді або сольових розчинах. Донедавна в цих білках розрізняли міогенну, альбумінову, глобулінову та міоглобінову фракції. Але ці фракції не однорідні. Так, міогенна фракція включає в себе ряд ферментів гліколізу, Неоднорідними є й інші білки саркоплазми. Зокрема тут виявлено білки-ферменти, що знаходяться в мітохондріях і відповідають за тканинне дихання. Міоальбумін саркоплазми за хімічними властивостями нагадує альбумін плазми крові, Міоглобін м’язів - типовий хромопротеїн, що, як і гемоглобін, з’єднується з киснем і забезпечує процес дихання м’язів. Червоний колір м’язів зумовлений великим вмістом у них міоглобіну. Міоглобін має в 5 разів більшу спорідненість із киснем, ніж гемоглобін. Це сприяє забезпеченню значного резерву кисню в м’язовій тканині при його нестачі.
Білки міофібрил. До складу міофібрил входять такі білки: міозин (56-60 %), актин (20-25%), тропоміозин (10-15 %) і тропоніновий комплекс (4-6 %).
Білки строми в поперечносмугастих м’язах представлені переважно колагеном, нейрокератином, еластином тощо. Ці білки входять до складу сполучнотканинних елементів стінок судин, нервів та сарколеми.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
Ліпіди. У м’язах знаходяться нейтральні жири, стериди, фосфоліліди. Нейтральні жири входять у простір між структурами м’язових волокон і відіграють роль резервного жиру. Їх вміст дуже непостійний.
Холестерин і фосфоліпіди є обов’язковими складовими компонентами всіх м’язів і входять до складу клітинних мембран. Вміст фосфоліпідів і холестерину в м’язах збільшується під час тренування.
Екстрактивні речовини м’язів. Скелетні м’язи містять ряд важливих екстрактивних речовин: нуклеотиди (АТФ, АДФ, АМФ, ТТФ, УТФ, ЦТФ, інозинмонофосфат), креатинфосфат, креатинін, карнозин, ансерин, карнітин тощо.
Серед них креатин та креатинфосфат мають пряме відношення до скорочення м’язів. В їх синтезі беруть участь 3 амінокислоти: аргінін, гліцин, метіонін. Утворення їх починається в нирках, а завершується в печінці і м’язах.
З амінокислот у м’язах найбільше глутамінової кислоти та глутаміну.
Безазотисті екстрактивні речовини м’язів представлені переважно вуглеводами та продуктами їх обміну. Найбільше в м’язах глікогену. У людини вміст глікогену в м’язах знаходиться в межах 0, 4-0, 8 %, але під впливом тренування він може збільшуватися до 1, 5-3%. Втомлені м’язи містять незначну кількість глікогену.
Під час роботи глікоген м’язів розпадається на глюкозу, тріозо-фосфорні ефіри та інші проміжні продукти гліколізу, в тому числі молочну кислоту.
Мінеральні речовини. Загальний вміст мінеральних речовин в м’язах на сиру масу становить 1, 0-1, 5 %. Із катіонів у м’язах переважають К+, Na+, Са2+, Мд2+, є також мідь, марганець, цинк; з аніонів - найбільше фосфатів та сульфатів. За рахунок іонів у м’язах підтримуються сталість рН і осмотична рівновага та здійснюється специфічний вплив на їх збудливість та скоротливість. Зниження концентрації солей у м’язах призводить до зменшення їх збудливості.
Структурною одиницею скелетного м’яза є багатоядерна клітина— міоцит— м’язове волокно, довжина якого у людини може досягати 10-12 см, а діаметр — 0, 01-0, 1 мм.
Міоцити оточені електрично збудливою плазматичною мембраною, що отримала назву сарколеми, містять у собі цитозольну частину — саркоплазму, скорочувальні елементи — міофібрили, високодиференційовану ендоплазматичну сітку — саркоплазматичний ретикулум, розвинену систему мітохондрій — саркосом (рис. 1).
В активно функціонуючих скелетних м’язах людини мітохондрії дуже чисельні; вони розміщуються впродовж міофібрил, у безпосередній близькості від них, що забезпечує мінімальну відстань для дифузії макроергічних фосфатів до скорочувальних елементів.
Скелетні м’язи мають розвинену систему мембран і канальців саркоплазматичного ретикулуму (СР), які відіграють роль головного резервуара внутрішньоклітинного Са2+ контролюючи активну концентрацію іонів кальцію в цитозолі. Зміни концентрації цитозольного Са2+ є біохімічним регулятором включення процесу м’язового скорочення. З елементами СР контактує система поперечних трубочок — Т-система, упродовж елементів якої відбувається поширення електричного потенціалу від сарколеми до мембран СР, спричиняючи вивільнення з СР іонів кальцію.
Значну частину об’єму м’язових клітин займають скорочувальні елементи — міофібрили, що спаковані в паралельні пучки. Молекулярною основою міофібрил є організовані у вигляді ниток (філаментів) скорочувальні білки — актин -міозин.
Саркомери — структурно-функціональні елементи скорочувального апарату скелетних м’язів. Саркомери утворені пучками міофібрил, які відокремлені один від одного перпендикулярними смугами — Z-лініями.
Електронно-мікроскопічне вивчення саркомерів дозволило виявити в них упорядковані елементи, які створюють характерну для скелетних м’язів поперечну смугастість (рис. 1.), а саме:
І-диски (ізотропні), що утворені " тонкими" філаментами (діаметром близько 6 нм) міофібрил;
А-диски (анізотропні), що утворені " товстими" філаментами (діаметром 15 17 нм) міофібрил, які перекриваються з " тонкими" філаментами;
Н-зона — частина А-диска, в якій " товсті" філаменти не перекриваються з " тонкими".
|

Рис. 1. Структурна організація саркомерів міофібрил.
До складу міофібрил входять такі білки:
1) до складу товстих ниток — білок міозин;
2) до складу тонких ниток — білки актин, тропоміозин, тропоніновий комплекс (тропонін Т, тропонін І, тропонін С);
3) білок а-октинін — компонент Z-лінії саркомерів; з цим білком сполучені кінці F-акгинових молекул тонких філаментів.
Міозин — фібрилярний білок, що утворює товсті філаменти міофібрил. Молекула міозину асиметрична, складається з двох важких поліпептидних ланцюгів, що мають конформацію 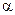 -спіралі й закручені один відносно одного; довжина молекули — 160 нм. N-кінці важких ланцюгів утворюють глобулярні " голівки", які нековалентними зв’язками сполучені з додатковими чотирма легкими поліпептидними ланцюгами. Глобулярна частина молекули міозину має дуже важливу особливість; в присутності іонів Са2+ вона проявляє АТФ-азну активність, тобто здатна розщеплювати аденозинтрифосфатну кислоту (АТФ), звільняючи тим самим хімічну енергію, необхідну для м’язового скорочення. Молекула міозину містить значну кількість глутамінової кислоти і тому має доситьзначнийнегативний заряд. Поперечні містки товстих виток - це глобулярні голівки білку міозину.
-спіралі й закручені один відносно одного; довжина молекули — 160 нм. N-кінці важких ланцюгів утворюють глобулярні " голівки", які нековалентними зв’язками сполучені з додатковими чотирма легкими поліпептидними ланцюгами. Глобулярна частина молекули міозину має дуже важливу особливість; в присутності іонів Са2+ вона проявляє АТФ-азну активність, тобто здатна розщеплювати аденозинтрифосфатну кислоту (АТФ), звільняючи тим самим хімічну енергію, необхідну для м’язового скорочення. Молекула міозину містить значну кількість глутамінової кислоти і тому має доситьзначнийнегативний заряд. Поперечні містки товстих виток - це глобулярні голівки білку міозину.
Фібрилярні " хвости" молекул міозину контактують між собою в поздовжньому напрямку, утворюючи товсті філаменти саркомерів, до складу кожного з яких входять, близько 400 молекул міозину. Глобулярні голівки виступають із зовнішньої поверхні філамента (рис. 3.).
|

Рис. 2. Будова молекули міозину
|

Рис. 3. Молекулярна організація товстого (міозинового) філамента
Актин — білок, що існує в двох формах: G- та F-актин. G-актин — глобулярний білок, що має вигляд кулеподібних молекул діаметром близько 5 нм. Молекули G-актину (субодиниці) нековалентно сполучаються між собою, утворюючи намистоподібні утворення — ланцюги фібрилярного F-актину.
У м’язових клітинах F-актин представлений фібрилярними структурами, що складаються з двох ланцюгів, переплетених один навколо одного. Особливістю G-актину є здатність дуже міцно зв’язувати один іон Са2+ і одну молекулу АТФ або АДФ.
При цьому зв’язування АТФ G-актином звичайно супроводжується його асоціацією з утворенням F-актину. Мономери актину, розщепляючи АТФ, здатні з’єднуватися в димери, які містять АДФ.
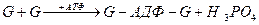
З димерів побудована подвійна спіраль полімерного фібрилярного актину.
У міофібрилах міститься також тропоміозин - білок, що утворює комплекс з білком тропоніном - глобулярним білком.Тропонін має значний негативний заряд. У розслабленому м’язі тропоміозин блокує нитки актину, а тропонін інгібує ферментативну активність міозину, запобігаючи розщепленню АТФ і звільненню енергії, необхідної для м’язового скорочення. Блокада знімається іонамиСа2+, які зв’язуються з комплексом тропонін-тропоміозин та інактивують його.
Тропоміозин — білкові молекули витягнутої форми, що складаються з двох поліпептидних ланцюгів ( та β), які утворюють подвійну спіраль. Паличкоподібні молекули тропоміозину (довжиною 40 нм і товщиною 2 нм) розміщуються в борозенках між двома ланцюгами F-актину таким чином, що кожна молекула тропоміозину контактує із сімома молекулами (субодиницями) G-актину.
Тропонін — білок тонких філаментів, що складається з трьох субодиниць: ТnТ, ТnІ, ТnС. Тропонінові комплекси мають глобулярну форму і розміщуються впродовж актинового філамента з інтервалами в 38, 5 нм, контактуючи з кінцями молекул тропоміозину (рис. 4.).
|
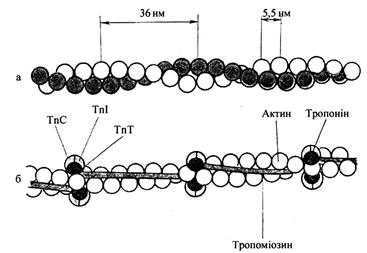
Рис. 4. Схема взаємодії тропоміозину (Tm) і тропонінів з актином: а — спіраль F-актину, б— взаємодія Тт та тропонінів із субодиницями G-актину.
В основу сучасного уявлення про механізми м’язового скорочення покладено запропоновану Г.Хакслі та іншими дослідниками модель ниток (філаментів), що плавно переміщаються (slide — awn.) впродовж одна одної. Головні постулати цієї моделі, що були підтверджені біохімічними, біофізичними й електронно-мікроскопічними дослідженнями:
- товсті(міозинові) та тонкі (актинові) філаменти міофібрил не змінюють своєї довжини протягом м’язового скорочення;
- протягом м’язового скорочення зменшується довжина всього саркомера внаслідок зустрічного руху та перекривання товстих і тонких філаментів;
- скорочувальна сила генерується внаслідок активної взаємодії одного типу філаментів з іншим, сусіднім типом філаментів.
Схема, що ілюструє взаємний рух товстих і тонких філаментів саркомера в процесі м’язового скорочення, подана на рис. 5.
Було також встановлено, що м’язове скорочення, в основі якого лежить переміщення товстих і тонких філаментів, потребує участі АТФ; циклічне перетворення АТФ в АДФ є необхідною передумовою як скорочення, так і розслаблення м’язів. Гідроліз АТФ до АДФ та Φ здійснюється завдяки АТФ-азній активності глобулярних голівок S1 міозину.
|

Рис. 5.Модель взаємного переміщення філаментів у саркомері
Залежна від АТФ взаємодія актину з міозином, що призводить до взаємного переміщення тонких і товстих філаментів саркомера, відбувається наступним чином (рис. 6.):
А – у м’язі, що перебуває в стані спокою, S1-голівки міозину не сполучені з актиновими філаментами. Продукти гідролізу АТФ (АДФ та Фн) зв’язані з міозином.
В – при збудженні м’яза S1-голівки зсуваються в напрямку тонких філаментів і сполучаються з нитками актину (G-субодиницями). Φ вивільняється з комплексу з міозином.
С – вивільнення АДФ з комплексу з міозином супроводжується конформаційним зсувом у просторовому розташуванні голівки S1, що зв’язана з актином (зміщенням кута між голівкою і віссю міофібрили з 90° на 45°).Зміна просторовоїорієнтації S1-голівки міозину відносно нитки актину призводить до розвитку напруги і пересування тонкого філамента відносно товстого приблизно на 100 А° (10 нм) у напрямку середини саркомера.
D – взаємодія з міозином молекули АТФ супроводжується розривом зв’язку між актином і міозином. S1-голівка знову віддаляється від тонкого філамента.
Е – АТФ, що вивільнився, гідролізується до АДФ та Φ, завдяки АТФ-азній активності вільних голівок міозину. Продукти гідролізу знову сполучаються з міозином. Актинові та міозинові філаменти готові до нового циклу взаємодії та пересування.
|

Рис. 6. Схема молекулярних механізмів м’язового скорочення
Загальна схема м’язового скорочення, що полягає у взаємному переміщенні актинових і міозинових філаментів, стосується усіх типів м’язів. Разом з тим, біохімічні механізми включення процесу та регуляція скорочення і розслаблення суттєво відрізняються в скелетному і серцевому м’язах (з одного боку) та гладенькому — з іншого.
Ініціація м’язового скорочення — процес, що включається генерацією потенціалу дії на сарколемі внаслідок хімічного сигналу, який надходить із нервово-м’язового синапсу (наприклад, при вивільненні холінергічним нейроном ацетилхоліну і взаємодії медіатора з локалізованим у сарколемі холіновим рецептором).
Скорочення м’язів - це зближення ниток актину і міозину. За соті долі секунди спрацьовує довгий ланцюг хімічних і фізико-хімічних процесів. Це відбувається за командою-імпульсом. Центральна нервова система посилає у м’язи електричні імпульси, що рухаються по нервах зі швидкістю 70-120 м/с. Проте до м’яза ці сигнали не доходять, тому що між нервовим закінченням і м’язовою клітиною немає контакту. Вони розділені так званою синаптичною щілиною, яка створює величезний опір електричному імпульсу. Щоб м’яз одержав команду, електричний сигнал має бути перетворений у хімічну форму. Перетворювачем електричного сигналу служить синапс.
Він складається з двох мембран - на закінченнях нерва (пресинаптична мембрана) і м’язового волокна (постсинаптична мембрана). Між двома мембранами знаходиться синаптична щілина. У пресинаптичній мембрані заховані сотні тисяч (до мільйона) пухирців з ацетилхоліном, який служить медіатором - хімічним передавачем сигналу.
Рис. 7. Схема будови нервово-м’язового синапсу
Під дією електричного імпульсу частина їх потрапляє в синаптичну щілину і досягає постсинаптичної мембрани. Цей медіатор і передає сигнал м’язовому волокну. Процес транссинаптичної передачі триває близько 5 * 10-4 с. Діючи на постсинаптичну мембрану (сарколему), ацетилхолін викликає її деполяризацію, з якої і починається ланцюг подій, що приводить до скорочення м’язового волокна. Деполяризація підвищує проникність клітинних мембран, у тому числі й мембран внутрішньоклітинних каналів, цистерн, в яких концентрація іонів Са2+ у 10000 разів вища, ніж у саркоплазмі. Збільшення проникності призводить до того, що іони Са2+’ " виливаються" із цистерн і концентрація їх у саркоплазмі зростає.
Іони Са2+ зв’язуються з комплексом тропонін-тропоміозин і інактивують його. У результаті звільняються заблоковані реакційноздатні ділянки актину і голівки молекул міозину. Маючи АТФ-азну активність, голівки міозину розщеплюють молекули АТФ, які вних знаходяться. У результаті звільненої енергії глобулярні голівки згинаються, прикріплюються до актинових молекул. При цьому утворюються поперечні міозинові містки, які, якгумки, тягнутьна себе молекули актину. Актинові нитки сковзають уздовж міозину, втягуються в щілини між міозиновими молекулами. М’язове волокно скорочується.
У міру використання енергетичних запасів кожної молекули АТФ зв’язок міозину з актином розривається, а на голівці утворюється нова молекула АТФ, гідроліз якої знову “заряджав” поперечний місток енергією. Цей процес повторюється багаторазово, приводячи до все більш глибокого просуванняактинових витокміж міозиновими молекулами. Так відбувається перетворення хімічної енергії безпосередньо в механічну.
У момент м’язового скорочення довжина товстих міозинових ниток залишається постійною. Не змінюється і довжина А диску. При помірному вкороченні не змінюється також і довжина тонких ниток, але зменшується довжина І-диску і саркомеру в цілому (відстань між Z-пластинками). На підставі цих даних і був зроблений висновок про те, що скорочення м’яза в цілому здійснюється шляхом ковзання тонких протофібрил (актину) у проміжках між товстими (міозином). У випадках надскорочення спостерігається наповзання тонких ниток у центрі саркомера одна на одну, а Z-пластинки надавлюють на кінці товстих протофібрил, Уцих випадках довжина їх дещо зменшується.
Поки в щілині е ацетилхолін, деполяризація м’язового волокна триває і воно не реагує на нові сигнали центральної нервової системи. Для нового скорочення необхідно зруйнувати медіатор. Робить це ацетилхолінестераза - фермент, який виділяє пресинтаптична мембрана. Після руйнування медіатора починається реполяризація, яка зменшує проникність цистерн. За допомогою так званого кальцієвого насоса іони Са2+ закачуються в цистерни проти дифузного градієнту а витратою енергії АТФ. Зникнення іонів Са2+ із саркоплазми звільняє тропонін, який інгібує АТФ-азну активність міозину і відповідно утворення енергії. Актин і міозин повертаються на свої місця. М’яз розслаблюється. Всі ці складні хімічні реакції мають високу швидкість. Через деякі синапси за 1 секунду може проходити до 300 команд імпульсів, тобто стільки разів виходять і руйнується ацетилхолін.
У відповідь на одиночний імпульс відбувається одиночне скорочення, яке супроводжується деполяризацією і реполяризацією м’язових волокон. Причому реполяризація відбувається до того, як м’яз повністю розслабиться. Якщо я цей момент надходить новий імпульс, скорочення ніби сумуються, саркомер (рухова одиниця) скорочується сильніше, ніж у випадку одиночного імпульсу. Відповідно росте і сила, яку розвиває м’яз. При частоті імпульсів 20-35 с-1 відбувається скорочення, яке у 2-3 рази переважає за силою одиночне.
За допомогою міографа можна виявити зубці окремих скорочень. Такий стан називають зубчатим тетанусом.
При подальшому збільшенні частоти імпульсів може настати такий стан, коли кожний новий сигнал буде припадати на майже повністю скорочений попереднім імпульсом м’яз. Тоді всі одиночні скорочення зливаються в одне. У цей момент м’язи розвивають найбільшу силу тяги, майже в 4 рази вищу, ніж при одиночному скороченні. Це гладкий тетанус.
У тому випадку, коли частота нервових імпульсів значно вища, ніж потрібно для гладкого тетануса, тоді через високу частоту виділення ацетилхоліну в синапсах і збільшення його концентрації холінестераза не встигає руйнувати його. М’язові волокна втрачають здатність сприймати нові імпульси, і сила рухових одиниць різко падає. Це явище одержало назву песимума. Воно спостерігається в стресових ситуаціях, у стані втомлення, при зусиллях, які перевищують фізіологічні можливості організму.
Література:
1. Биохимия: Учебник / Под ред. В.В. Меньшикова, Н.И. Волкова. -М., Физкультура и спорт, 1986. - 384 с.
2. Біохімія: Посібник / Під ред. О.Ф. Явоненко, В.В. Яковенко. – Суми, Університетська книга, 2002. – 379 с.
3. Биохимия мышечной деятельности и физической тренировки: Учебник / Под ред. Р.Мохан, М. Глессон, П. Гринхафф. – Київ, Олимпийская литература, 2001, - 295 с.
|
|