Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Методи культивування вірусів та їх оцінка. Виявлення для репродукції вірусів в курячих ембріонах та культурі клітин. Реакція гемаглютинації, цитопатична дія вірусів, її види.
|
|
І. у цілому живому організмі (рослин, тварин) - це був перший метод. Оцінка достоїнств: 1) вивчаємо взаємодію вірусів із цілим живим організмом; 2) чітко бачимо результат; 3) можемо вивчити розвиток хвороби - патогенез;
4) випробуємо вакцини, способи (препарати) лікування; 5) накопичуємо велику кількість вірусів Недоліки: 1) в організмі є свої бактерії і віруси, 2) не стерильна система для одержання вакцин; 3) дорогий метод; 4) не універсальний метод - не можна знайти чутливих тварин
II. у 30-ті роки Бернет запропонував метод культивування в куриному ембріоні (КЕ).Оцінка достоїнств1) стандартний - використовують 9-1 2 денні КЕ;
2) стерильність; 3) можемо заражати різні порожнини (жовточний мішок, ембріон, алантоісна порожнина); 4) більш економічно.Недоліки: 1) не всі віруси можна культивувати в КЕ; 2) у КЕ можуть бути віруси лейкозу курок.
ІІІ. у культурі клітин (КК) — це сукупність клітин, отриманих із цілого живого організму, які культивуються іп Vitro (поза організмом) Оцінка достоїнств: 1) вивчаємо взаємодію вірусу з клітинами,
2) можна одержувати вакцини; 3) багато вірусів можна культивувати
Недоліки: 1) дуже складний і дорогий метод
Принцип одержання культури клітин:
1) із живого організму одержують шматочок тканини (печінки, нирки, пухлини, ембріона), 2) шматочки роздрібнюють, а потім за допомогою трипсина розділяють до окремих клітин; 3) клітини поміщають у спеціальне живильне середовище і помішають у камеру Горячева для підрахунку клітин, 4) клітини в середовищі поміщають у термостат (37°С для росту).Умови для культивування клітин1) стерильність (посуд, повітря),
2) повноцінне живильне середовище - використовують дуже складні середовища, що містять білки, вуглеводи, ліпіди, амінокислоти, вітаміни, мінеральні солі. Для стерилізації в них добавляють антибіотики.
Виявлення вірусу: ІІ. у куриному ембріоні: 1) по загибелі ембріона;
2) відставання (або затримка в розвитку); 3) по крововиливах і осередках некрозу на хорион алатоїсной оболонці; 4) по реакції гемаглютинації (РГА) - це склеювання еритроцитів під впливом вірусів, які мають гемаглютинуючі властивості. Механізм РГА: на поверхні деяких складних вірусів є особливі рецептори - гемаглютиніни. Віруси адсорбуються на поверхню еритроцитів, завдяки гемаглютинінам вони викликають склеювання еритроцитів і утворення осаду з нерівним краєм. У нормі еритроцити осідають, характер осаду - рівний край. Оцінка достоїнств РГА: 1) РГА чітко видно в порівнянні з контролем; 2) проходить швидко (1 година) у нестерильних умовах;
3) залежить від виду вірусів (віруси грипу, подагри, віспи) та від виду еритроцитів і (наприклад, вірус грипу склеює еритроцити людини і курок).
Недоліки РГА: 1) не універсальна реакція (не всі віруси мають гемаглютиніни).
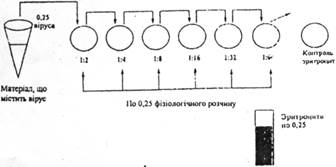
Висновок: РГА пройшла, отже, ми виявили вірус, який мас гемаглютинуючі властивості. Титр вірусів у РГА 1/32 - це найбільше розведення вірусу, що викликає РГА.
ІІІ. В культурах клітин: І) по кольоровій індикаторній реакції - живі клітини змінюють рН і колір індикатора, а мертві не змінюють; 2) по цитопатогенній дії вірусу (ЦПД) - спостерігаємо при малому збільшенні мікроскопа.
Види ЦПД: A) повна деструкція (руйнація) клітин — наприклад, ситеровіруси, B) часткова деструкція: a) осередкова; b) гроноподібна - аденовіруси; c) утворення симпластів або синцитіїв - це злиття уражених клітин між собою (наприклад, віруси кору, паротиту, ВІЛ, респіраторно синтиціальний вірус RS).
C) проліферації - це необмежене розмноження клітин - онкогенні (пухлинні) віруси.
3) по утворенню бляшок - це зона руйнації клітин під дією одного вірусу (умовно). Для одержання бляшок культуру покривають тонким шаром агара або бентоніта (бенгоніт — це глина, має властивості геля, її застосування для вірусних бляшок вивчив В.П.Широбоков. Гель мішає поширенню вірусів, тому утворюються окремі зони загибелі клітин - бляшки). Метод якісний і кількісний;
4) по реакції гемадсорбції — РГ адсорбції — це адсорбція еритроцитів на поверхню клітин, уражених вірусами (гемадсорбуючими).
Механізм РГ адсорбції: Складні віруси при виході з клітини змінюють її мембрану - відбувається відбруньковування. До цих змінених місць можуть прикріплюватися еритроцит (наприклад, парамексовіруси).
5) по утворенню вірусних включень — це ділянки клітини, у яких відбувається обмеження репродукції вірусів. Включення забарвлюються інакше, ніж цитоплазма (наприклад, включення Бабеша-Негрі в нейронах при сказі).
Зараження клітинних культур: 1) узяття матеріалу, що містить вірус (кров і т.ін.);
2) підготування матеріалу: a) профільтрувати через бактеріальний фільтр;
b) додати антибіотики - пеніцилін і стрептоміцин.3) зараження клітинної культури:
a) вилити середовище росту; b) нанести на клітини 0, 1 мл вірусу і залишити для адсорбції на 15-20 хвилин; c) внести в пробірки середовище підтримки. В дослідній пробірці був вірус, у контрольній - ні.
Зараження куриного ембріона: 1) закритий спосіб - та допомогою шприца в різні місця — у хорион аллантоїсну порожнину, у жовточний мішок, в ембріон, на хорион аллантоїсну оболонку; 2) відкритий спосіб -- роблять отвір, проводять зараження, закривають склом (покривним і парафіном), спостерігають зміни.
64. Серологічні реакції у вірусології. PH, РГГА, РЗК... Серологічні реакції для виявлення вірусів: PH, РГГА, РЗК, кольорова проба (вірус знищує клітини і через відсутність метаболізму колір середовища залишається незмінним), РІФ. PH - антитіла імунної сироватки здатні нейтралізувати ушкоджуючу дію вірусів на чутливі клітини тканин. Відповідність вірусу використаній антисироватці виявляється придушенням бляшкоутворення (із застосуванням агарового чи бентонітового (Широбоков) покриття) у порівнянні з контролем. РГГА - заснована на здатності антитіл імунної сироватки нейтралізувати віруси, що у результаті цього процесу утрачають властивості аглютинувати еритроцити. Застосовують для діагностики грипу, кору, краснухи, кліщ енцефаліту й інших вірусів які можуть аглютинувати еритроцити різних тварин. РЗК - 1 фаза інкубації суміші шуканого антигену з діагностичною сироваткою і комплементом; 2 фаза - індикаторна - визначення вільного комплементу додаванням гемолітичної системи (еритроцити барана і гемолітична сироватка утримуюча антитіла до еритроцитів барана). При негативній реакції, якщо антиген і антитіло не відповідають один одному, комплемент, залишається вільним і в 2 фазі реакції він приєднується до комплексу эритроцит - антиэритроцитарное ##### гемолітичної сироватки, викликаючи гемоліз. Використовується для ретроспективної діагностики багатьох вірусних інфекцій на підставі виявлення специфічних антитіл у парних сироватках крові людей, а також для визначення вірусоспецифічних антигенів у різних матеріалах отриманих від хворого.
65. Сучасні методи Твердофазний імуноферментний аналіз. Належить до основних методів виявлення антитіл окремих класів. Існують різні модифікації методу, але в основі тест-систем для виявлення антитіл є відповідні антигени, адсорбовані на " твердій фазі" - на стінках лунок полістиролових планшетів, на полістиролових булавкоподібних кульках з тримачами, на хроматографічних смужках - стріпах та ін. Антитіла досліджуваної сироватки з'єднуються з цими антигенами і таким способом також фіксуються на " твердій фазі" і не вимиваються промивним розчином. Ці антитіла виявляють за допомогою мічених ферментом сироваток проти окремих класів імуноглобулінів людини. Далі виявляють фермент, що фіксувався на твердій фазі в результаті реакцій між міченою сироваткою і імуноглобуліном. Для цього в систему вносять субстрат для ферменту і хромоген - речовину, яка змінює колір внаслідок ферментативної реакції. Облік реакцій ведуть в автоматичному режимі за допомогою спеціальних фотометрів - рідерів, хоча досить часто застосовують візуальний облік, порівнюючи інтенсивність забарвлення хромогену із стандартною шкалою. Схема реакції: вірусний антиген (на твердій фазі) + досліджувана сироватка, промивання + мічена ферментом сироватка проти Lg людини, промивання + субстрат для ферменту і хромоген. Зміна кольору хромогену свідчить про позитивну реакцію. Найчастіше використовують сироватки, мічені пероксидазою, тоді субстратом є пероксид водню, а хромогеном ортофенілендіамін. Впровадження ІФА значно спростило дослідження, підвищило його достовірність і доступність для практичних лабораторій.
Полімеразна ланцюгова реакція (ПЛР). Належить до останнього покоління діагностичних методів і широко застосовується, крім мікробіології та вірусології, в багатьох галузях біології та медицини, зокрема при судово-медичній експертизі. Метод ґрунтується на " розмноженні" - ампфілікації і виявленні окремих специфічних генів. Відомо, що синтез молекул ДНК в клітині відбувається за матричним принципом - на одній з ниток ДНК за допомогою ферменту ДНК-полімерази за правилом компліментарності синтезується друга нитка. Реакція відтворюється і пробірці, для цього необхідна ДНК-матриця, компоненти нуклеїнової кислоти - нуклео-тиди і ДНК-полімераза. Але ДНК-полімераза починає синтез тільки у тій точці, де вже є двоспіральна ділянка молекули -" затравка". Для ПЛР необхідні праймери - синтетичні олігонуклеотиди, що за своєю послідовністю відповідають специфічним ділянкам розміщеним на кінцях гена, який виявляється. Реакція відбувається у спеціальних автоматичних приладах - ампфілікаторах. ДНК з досліджуваного матеріалу нагрівається, при цьому вона розкручується і в систему додають праймери. Праймери приєднуються до специфічних ділянок досліджуваного гена, температура в системі знижується, і ДНК-полімераза синтезує копію гена. Далі цикл повторюється - нагрівання, реакція з праймерами, синтез нових копій гена. Протягом 30- 40 циклів синтезуються мільйони копій гена, які потім легко виявити за допомогою реакції молекулярної гібридизації. Зрозуміло, що метод розроблений на останніх досягненнях молекулярної біології, генетики та біохімії, завдяки яким стало можливим виділити окремі гени, розшифрувати послідовність нуклеотидів у них і синтезувати праймери.
66. Використання клітинних культур у вірусології. Класифікація культур клітин. Поживні середовища для культивування клітин. Класифікація клітинних культур Враховують такі ознаки: кількість пасажей (перенос клітин у нове середовище) і походження клітин.
Типи клітинних культур:
1. Первинно трипсинізовані - одержують з швидкозростаючих ембріональних тканин (людини, мавпи), кіл-сть пасажей 8-10;
2. Пасажні або перещеплювані – одержують з пухлинних тканин, кіл-сть пасажей не обмежено. Клітини злоякісні. Не можливо вивчати взаємодію з вірусом. Але для виділення вірусів ці кліт зручні.
3. Напівперещеплювані або диплоїдні клітинні штами - одержують з норм кліток, набір хромосом диплоїдний,, кіл-сть пасажів до 100.Використ для одержання вірусних вакцин. Це клітини шкіри, легенів.
- Живильні середовища для культур кліток містять повний набір амінокислот, вітамінів, ростові фактори: гідролізат лактальбуміна, сухі середовища і концентрати, які перед використанням розводять. Для вирощування клітинних культур використовують ростові середовища, збагачені сироватками тварин і людини. (бичача сироватка, ембріональна коров'яча сироватка)
67. Види взаємодії вірусів і клітин. Віруси є облігатними внутрішньоклітинними паразитами. Вони проходять всі основні етапи репродукції (розмноження). За кінцевим результатом взаємодії вірусу з клітинами виділяють такі типи:
I. продуктивний тіш взаємодії - характеризується тим, що в результаті проникнення і репродукції клітина гине, з неї виходить багато вірусів (вірусне потомство);
II. інтегративний — характеризується тим, що відбувається об'єднання або інтеграція генома вірусу і клітини. Найбільш характерний такий тип для ДНК геномних вірусів (герпеса, аденовірусів), для РНК геномних інтеграція можлива за допомогою ферментів оберненої транскриптази (ОТ). ОТ - переписує інформацію з РНК вірусу, синтезує його ДНК-копію. Вона інтегрує з генома клітини. Вірус у такому стані називається провірусом (вірус імунодефіциту людини — ВІЛ).
Усі властивості вірусів як біологічних об'єктів та патогенних агентів проявляються при їхній взаємодії з чутливим клітинами. Ця взаємодія відбувається за кількома стадіями:
1. Специфічна адсорбція віруса на клітині внаслідок взаємодії білків віріона і особливих рецепторів клітини.
2. Проникнення віруса в клітину (віропексис) відбувається шляхом захоплення віріона клітиною (піноцитоз).
3. Депротеїнізація віріона - звільнення вірусного геному від білкових оболонок - " роздягання". Здійснюється при злитті оболонки віруса з мембранами клітини та при дії клітинних ферментів.
4. Синтез вірусних компонентів. Цю стадію називають " екліпс-фазою", фазою " затемнення" - в цей час вірус не виявляється як індивідуальна структура або інфекційний агент. Екліпс-фаза властива тільки вірусам.
Під час вказаної фази здійснюються процеси транскрипції - синтезу вірусних матричних молекул мРНК для ДНК-вмісних вірусів або плюс-РНК для вірусів, що мають РНК мінус-типу. З участю інформаційних молекул відбувається синтез вірусних білків на рибосомах клітини (трансляція). Одночасно йде відтворення вірусного геному - синтез віріонних ДНК або РНК (редуплікація). Залежно від типу вірусів, процеси транскрипції і редуплікації відбуваються у ядрі або цитоплазмі клітини, а трансляція - у цитоплазмі.
5. Утворення нових вірусних часток. У простих вірусів віріони утворюються шляхом самозбирання. Нуклеокапсиди складних вірусів утворюються також при самозбиранні, а суперкапсидна оболонка
формується з участю мембран клітини. Ліпіди суперкапсидної оболонки — клітинного походження, а білки - вірусного.
6. Вихід віруса з клітини. Може відбуватися одномоментно, клітина при цьому руйнується, або поступово - віріони наче " відшнуровуються" від поверхні клітини. Але через деякий час клітина також гине.
68 Неспецифічні фактори захисту Інтерферони. Особливе місце в системі противірусного захисту відіграють білки-інтерферони. Ці білки утворюються клітинами (лейкоцити, макрофаги, фібробласти та ін.) у відповідь на вірусну інфекцію. Інтерферони не діють на вірус поза клітинами, але припиняють репродукцію віруса в клітині. Для них характерний ефект післядії - оброблена інтерфероном клітина стає резистентною до вірусів, навіть якщо інтерферон видалено. Дія інтерферонів неспецифічна - інтерферон, що утворився при дії одного віруса припиняє репродукцію будь-якого іншого віруса. Проте інтерферони видоспецифічні - клітини людини захищає тільки людський інтерферон. Дія інтерферонів не обмежується противірусною активністю. Вони мають протипухлинний, імуномодулюючий та інші ефекти Одержано лікувальні препарати інтерферонів. Найефективнішими є препарати генноінженерних (рекомбінантних) інтерферонів, які можна застосовувати в достатніх дозах парентерально. Препарати лейкоцитарного інтерферону застосовують місцево..
Специфічний противірусний імунітет. Здійснюється завдяки синтезу специфічних антитіл (гуморальний механізм) та специфічних імунокомпетентних клітин (клітинні механізми).
Антитіла нейтралізують віруси поза клітинами, але не впливають на репродукцію вірусів у клітинах. Слизові оболонки захищають секреторні імуноглобуліни класу А - sLgА.
Клітинний імунітет при вірусних інфекціях спрямований на клітини, в яких розмножується вірус. Під час репродукції вірусні антигени вбудовуються в зовнішню мембрану клітини, де розпізнаються імунокомпетентними клітинами. Таке розпізнавання можливе, лише якщо на поверхні клітини є антигени головного комплексу сумісності (НLА) - " подвійне розпізнавання". Деякі віруси мають механізм пригнічення антигенів НLА, ушкоджені ними клітини не розпізнаються імунною системою, що спостерігається при смертельних повільних вірусних інфекціях.
Надмірна реакція імунної системи на ушкоджені вірусом клітини може призвести до масової загибелі клітин, порушення функції органів і навіть до смертельних наслідків, наприклад при тяжких формах гепатиту В.
69. Вірусні вакцини Вакцини-препарати що отримані з мікробів продуктів їх життєдіяльності їх синтетичних або генно-інженерних аналогів які використовуються для активної імунізації людей та тварин з метою специфічної профілактики та терапії Класифікація за походженням: 1.корспускулярні (з цілих мікробних клітин): живі, вбиті. 2. вакцини з антигенних комплексів: хімічні. Субодиничні. 3. Анатоксини. 4. синтетичні. 5. генно-інженерні 6. антиідіотипічні Контроль: на стерильність на відсутність шкідливості на специфічну активність (імуногенність) на реактогенність (на лаб твар або добровольцях) на онкогенність Подвійний контроль (в спец інституті та на заводі)
Вірусні живі вакцини: віспяна (на основі вірусу віспи корів); корева (зі штаму Л-16 і штаму Едмонстон, отриманих А. А. Смородінцевим і М. П. Чумаковим); поліомієлітна (зі штамів А. Себіна типів 1, 2, 3); проти жовтої лихоманки (зі штаму 17D); грипозна (з лабораторних штамів, отриманих В. М. Ждановим і ін.); проти венесуельського енцефаломієліта коней (зі штаму 230, отриманого В. А. Андрєєв і А. А. Воробйов); паротитна (зі штамів, отриманих А. А. Смородінцевим і Н. С. Клячко).
Ортоміксовіруси. Історія відкриття, біологічні властивості, класифікація. Методи лаб діагностики грипу та їх оцінка. Проблема специфічної профілактики і терапії грипу. Лік-проф препарати та їх оцінка.
Довгий час збудником грипу вважали паличку, дрібну Гр.-бактерію. Вірус грипу відкритий Афанасьєвим і Фейфером. Цей вірус назвали також збудником інфлюенци (грипу). У 1933р Сміт і індус виділили з носоглотки хворих грипом вірус, що назвали вірусом грипу А. У 1940р Ференсис і Меджил відкрили вірус грипу, що відрізнявся від першого. Знову відкритий вірус назвали вірусом грипу В. У 1947р Тейлор виділив вірус грипу С. Потім були відкриті підтипи або варіанти вірусу грипу А-А0, А1, А2. До родини Orthomyxoviridae (orthos - правильний, myxa - слиз) належать віруси грипу А, В, С. Вірус грипу має сферичну форму, його розміри 80-120 нм. Геном утворений однонитковою РНК, яка складається з восьми фрагментів. Вона оточена білковим капсидом. Нуклеокапсид має спіральний тип симетрії. На зовнішній, суперкапсидній оболонці у вигляді шипиків розміщені гемаглютинін (Н) і нейрамінідаза (N). Гемаглютинін забезпечує адгезію вірусу на епітеліальних клітинах верхніх дихальних шляхів і його проникнення в цитоплазму. Нейрамінідаза має ферментативні властивості й сприяє виділенню новоутворених вірусів із клітини. Обидва глікопротеїни (N i H) мають виражені антигенні властивості. За внутрішнім нуклеопротеїдним антигеном розрізняють три типи вірусів грипу - А, В, С, які можна визначити в РЗК. У вірусу типу А людини є три різновидності гемаглютиніну (Н1, Н2, Н3) і дві - нейрамінідази (N1, N2). Залежно від їх комбінацій, виділяють підтипи вірусів
у А H1N1, H2N2, H3N2. Їх визначають у реакції гальмування гемаглютинації (РГГА) з відповідними сироватками. Вірус грипу А дуже мінливий. У середньому ч/з 10 років з’являється новий антигенний варіант, що призводить до виникнення чергової глобальної епідемії захв. Віруси грипу легко культивуються в курячих ембріонах і різноманітних культурах клітин. Максимальне накопичення вірусів відбувається ч/з 2-3 дні. У зовнішньому середов. вірус швидко втрачає інфекційність ч/з висушування. При низькій t в холодильнику зберігається протягом тижня, при -70° С - значно довше. Нагрівання призводить до його інактивації ч/з кілька хвилин. Під впливом ефіру, фенолу, формаліну швидко руйнується. Грип характеризується масовістю захворювань, які можуть охопити 10-50 % населення. Такі спалахи грипу А виникають кожні 1-2 роки в зимовий період. Епідемії грипу В реєструються кожні 3-4 роки. Резервуаром вірусу є хворі люди і вірусоносії. Проте в природі віруси можуть тривалий час перебувати в організмі свійських і диких тварин і птахів. Людина заражається повітряно-краплинним шляхом. Інкубаційний період триває 1-2 дні. Віруси перебувають у носоглотці за 1-2 дні до початку хв. і стільки ж після появи перших симптомів. У цей час людина найбільш заразна при чханні, кашлі, розмові. Вірус може проникати в кров і розноситись по тканинах і органах. У патогенезі захв. важливу роль відіграє інтоксикація й дія нейрамінідази, яка зменшує в’язкість слизової плівки, оголюючи рецептори епітеліальних клітин. Це сприяє розповсюдженню вірусів у трахею й бронхи, ушкодженню миготливого епітелію, проникненню вторинної мікрофлори - стафілококів, стрептококів, пневмококів. клебсієл та ін. В такому разі виникають такі ускладнення як пневмонія, бронхіт, плеврит, отит, менінгіт, енцефаліт. Важливу роль в постінфекційному імунітеті відіграють SIgA, інтерферон, антитіла проти гемаглютиніна й нейрамінідази. Несприйнятливість до вірусу грипу типу А триває 1-2 роки, типу В - 3-5 років, типу С - протягом усього життя. Лабор. діагностика проводиться з метою виділення вірусів шляхом зараж. змивом із носоглотки курячих ембріонів або клітинних культур та їх ідентифікації за допомогою РІФ, РГГА, РН з використанням специфічних протигрипозних сироваток. Для експрес-діагностики використовують ІФА, РІФ. Серологічні дослідження проводять за методом парних сироваток у РЗК, РГГА, ІФА та ін. Для специфічної профіл. грипу виготовлені різноманітні інактивовані вакцини: віріонні, субодиничні та їх комбінації. Застосовують живі вакцини. Проте їх ефективність досить низька. Для пасивної профіл. використовують протигрипозний Ig. Із лікувальною метою застосовують інтерферон, ремантадин, а при бактеріальних ускладненнях - АБ й сульфаніламідні препарати.
|
|