Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника

Единицы транскрипции (транскриптоны)
|
|
Синтез РНК молекулами РНК-полимераз in vivo начинается в определенных местах ДНК, называемых промоторами, и завершается на особых регуляторных последовательностях – терминаторах. Последовательности нуклеотидов ДНК, заключенные между промоторами и терминаторами, называют транскрипционными единицами, или транскриптонами. В пределах каждого транскриптона транскрибируется только одна цепь ДНК, которая получила название значащей или матричной. Термины " транскрипционная единица" или " транскриптон" по смыслу близки термину " ген", но они не всегда совпадают. Так, транскрипционные единицы прокариот, как правило, заключают в себе генетическую информацию нескольких генов и называются оперонами. Продуктами транскрипции оперонов являются полицистронные мРНК, в результате трансляции которых рибосомами образуется несколько белков. Белки, кодируемые полицистронными мРНК, обычно функционально связаны друг с другом и обеспечивают протекание какого-либо метаболического процесса, например биосинтеза определенной аминокислоты или утилизацию углеводов в качестве источника углерода. Организация генов в виде оперонов облегчает координированную регуляцию их экспрессии на уровне транскрипции. Согласованная регуляция транскрипции (и других этапов экспрессии) многих генов, не образующих одного оперона, чаще всего осуществляется специфическими белками-регуляторами, которые взаимодействуют с гомологичными регуляторными нуклеотидными последовательностями, маркирующими гены данной группы.
Промоторы эубактерий. В 1964 г. Ф. Жакобом, А. Уллманом и Ж. Моно была впервые сформулирована концепция промотора как особого регуляторного элемента в составе lac -оперона E. coli. Вскоре после этого были описаны промоторные мутации и высказано предположение, что промоторные участки оперона должны распознаваться молекулами РНК-полимеразы, а также заключать в себе сайт инициации транскрипции. В соответствии с генетическими критериями промотором может называться последовательность нуклеотидов, мутации в которой характеризуются следующими свойствами: 1) оказывают влияние на транскрипцию всех генов оперона; 2) действуют в цис-, но не транс- положении по отношению к генам, контролируемым мутантными промоторами; 3) локализуются вблизи одного из концов оперона, однако отличаются от мутаций в структурных частях генов тем, что не оказывают влияния на активность кодируемых этими генами ферментов. В соответствии с биохимическими критериями промотор представляет собой последовательность нуклеотидов, обеспечивающую базальный (но не максимальный) уровень транскрипции соответствующего транскриптона. Он является той минимальной последовательностью, которая специфически распознается холоферментом РНК-полимеразы среди случайных последовательностей нуклеотидов, что обнаруживается по образованию в определенных условиях стабильных комплексов фермент–ДНК.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
Структура обобщенного минимального промотора для холофермента РНК-полимеразы (Es70) эубактерий схематически представлена на рис. I.4, б. Такие промоторы содержат две канонические последовательности: область ‑ 35, обычно расположенную между нуклеотидами в положениях –30 и –35, и область –10, локализованную между нуклеотидами –7 и –10 (знак " минус" указывает на то, что нуклеотиды находятся перед первым транскрибируемым нуклеотидом, которому соответствует положение +1). Обе эти последовательности специфически взаимодействуют с двумя участками полипептидной цепи s70, называемыми s4.2 и s2.4. Расстояние между обсуждаемыми последовательностями оказывает влияние на активность промотора. Длина отрезка в 17 п.о. является оптимальной. Среди других последовательностей, оказывающих влияние на активность промоторов, но не являющихся универсальными, следует упомянуть так называемый UP-элемент – АТ-богатую последовательность, центр которой находится в положении –52 у промотора P1гена rrnB, кодирующего рРНК. С этим участком взаимодействует a-субъединица РНК-полимеразы. У нескольких других промоторов был обнаружен TG-элемент (центр – в положении –15 или –14), мутации в котором снижают активность этих промоторов.
Влияние мутаций в областях –10 и –35 на активность промотора, как правило, хорошо коррелирует с тем, насколько новая мутантная последовательность соответствует канонической. Однако в ряде случаев наблюдаются отклонения от этого правила, что позволило идентифицировать последовательности, важные для функционирования этих промоторов in vivo, с центрами в положениях –43 (область USR – upstream region), а также между нуклеотидами в положениях –1 и +20 (область DSR – downstream region). Эти последовательности не требуются для узнавания промотора РНК-полимеразой, однако облегчают процесс освобождения промотора после инициации транскрипции (см. ниже). Все описанные последовательности обеспечивают базальный уровень функционирования бактериальных промоторов. Однако активность промоторов может изменяться под действием многочисленных регуляторных факторов, взаимодействующих с другими регуляторными последовательностями, расположенными перед такими промоторами. Эти аспекты регуляции активности промоторов будут подробно рассмотрены в разделе 3.1.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
Промоторы эукариот. Промоторы эубактерий были определены выше как минимальный набор последовательностей нуклеотидов, необходимых для их специфического распознавания холоферментом РНК-полимеразы и начала (инициации) синтеза РНК. Распространение этого определения на промоторы эукариот встречает трудности из-за невозможности в настоящее время исчерпывающе описать эукариотический функциональный аналог бактериального холофермента Es.
Промоторы РНК-полимеразы II. Промоторы, узнаваемые РНК-полимеразой II, содержат три различных семейства регуляторных последовательностей ДНК. Последовательности первого семейства включают так называемые к ó ровые, или базальные элементы промотора, расположенные вблизи точки инициации транскрипции (см. рис. I.4, б). В настоящее время известны два класса базальных элементов: TATA-последовательность, расположенная за 25–30 нуклеотидов до точки инициации (каноническая последовательность – TATAa/tAa/t), и так называемый инициатор (Inr), последовательность которого обогащена пиримидинами (каноническая последовательность – PyPyA+1NT/APyPy). В этих обозначениях строчные буквы соответствуют нуклеотидам, которые присутствуют не во всех последовательностях, Py – пиримидиновые нуклеотиды инициатора, N – любой нуклеотид. Подстрочная цифра указывает на то, что с данного нуклеотида начинается транскрипция (точка инициации транскрипции). Элементы TATA-последовательности и инициатор необходимы для сборки ДНК-белкового инициационного комплекса и распознаются основными факторами транскрипции. Промоторы РНК-полимеразы II содержат один или оба регуляторных элемента или же не имеют их вообще. При этом оба элемента могут функционировать независимо друг от друга или же в их действии наблюдается синергизм.
К двум другим классам цис- регуляторных промоторных элементов у эукариот относятся последовательности, расположенные вблизи промотора (от 50 до нескольких сотен пар оснований перед точкой инициации), а также дистальные элементы (энхансеры и сайленсеры), расстояние которых от промотора может превышать 60 т.п.о. Оба класса таких последовательностей содержат сайты связывания регуляторных белков, модулирующих транскрипцию.
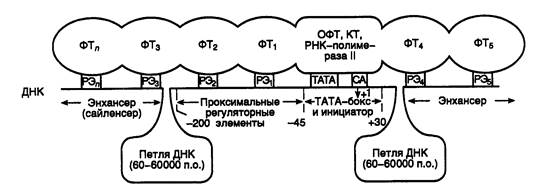
Рис. I.5. Межмолекулярные взаимодействия на промоторе РНК-полимеразы II
РЭ1–РЭn – последовательности нуклеотидов регуляторных элементов промотора, ФТ1–ФТn – взаимодействующие с ними регуляторные факторы транскрипции, ОФТ – основные факторы транскрипции, КТ – коактиваторы транскрипции, +1 – точка инициации транскрипции
Обобщенная схема промотора РНК-полимеразы II в составе инициационного комплекса представлена на рис. I.5. На ней отображены многочисленные белок–белковые и белково–нуклеиновые взаимодействия при инициации транскрипции. На этой схеме также указано наличие во многих промоторах консервативной последовательности CA, которая находится непосредственно перед нуклеотидом А+1. У некоторых промоторов, в частности ассоциированных с генами домашнего хозяйства, может отсутствовать явно выраженная TATA-последовательность. Проксимальные и дистальные регуляторные элементы промоторов построены из коротких транскрипционных элементов длиной в 10–15 п.о., с которыми непосредственно взаимодействуют факторы транскрипции. Проксимальные регуляторные элементы, как правило, имеют простую структуру, включающую один или несколько транскрипционных элементов (ТЭ). В то же время энхансеры устроены более сложно. Следует заметить, что в активном промоторе дистальные и проксимальные регуляторные элементы сближены друг с другом, в результате чего происходит " выпетливание" разделяющих их протяженных последовательностей ДНК.
Промоторы РНК-полимеразы III. Как уже упоминалось выше, гены, транскрибируемые РНК-полимеразой III, можно разделить на три основных класса (см. рис. I.4, б). К 1-му классу относят гены 5S рРНК, которые содержат внутреннюю регуляторную область (internal control region – ICR). Три элемента ICR, с которыми взаимодействует основной фактор транскрипции TFIIIA, локализованы между нуклеотидами +50 и +64. Комплекс TFIIIA–ICR стабилизируется после взаимодействия с TFIIIA фактора TFIIIC, хотя для последнего в промоторе отсутствует специфический участок связывания (подробнее о факторах транскрипции см. в разделах 2.1.3 и 3.2). Мутации, локализованные в окрестностях точки инициации транскрипции промоторов классов 1 и 2, оказывают сильное влияние на транскрипцию этих генов, что указывает на возможное присутствие в этих промоторах дополнительных регуляторных элементов.
Гены тРНК относят ко второму классу. Для промоторов этих генов характерно наличие внутренних блоков регуляторных последовательностей A и B. Блок А локализован между нуклеотидами +10 и +20, тогда как блок B удален от него на расстояние 30–60 п.о. по направлению к 3’-концу гена. Эти регуляторные элементы узнаются непосредственно фактором TFIIIC, который образует с ними очень прочный комплекс. По крайней мере, четыре полипептидные цепи, входящие в состав комплекса TFIIIC–промотор, контактируют друг с другом и покрывают участок ДНК между положениями –25 и +75. Канонический регуляторный элемент в окрестностях точки инициации транскрипции был локализован путем анализа последовательностей нуклеотидов в соответствующих генах тРНК.
Гены третьего класса кодируют малые ядерные РНК, например U6-РНК у высших эукариот, и содержат только регуляторные элементы, расположенные перед точкой инициации транскрипции. У большинства генов этого класса регуляторные элементы состоят из ТАТА-последовательности с центром в положении –30 и из проксимального элемента PSE (proximal sequence element) с центром в положении –60. Несмотря на наличие ТАТА-элемента, обычно определяющего положение точки инициации транскрипции, в данном случае именно PSE фиксирует положение этой точки. Аналогичную функцию у промоторов РНК-полимеразы II выполняет ТАТА-последовательность. Состав белковых компонентов, образующих комплекс с PSE, в настоящее время до конца не выяснен. Обнаружены два различных PSE-связывающих белковых комплекса, названные SNAP и PTF. Активность промоторов третьего класса зависит от наличия ТАТА-связывающего белка (TBP). Белок ТВР, входящий в состав фактора транскрипции TFIIIB, взаимодействует непосредственно с ТАТА-элементом промотора. Поскольку для инициации транскрипции на этих промоторах требуется наличие PSE-элемента, предполагают, что PSE-связывающий фактор взаимодействует с ТВР-компонентом фактора TFIIIB. Интересно, что промотор гена U6-РНК животных, узнаваемый РНК-полимеразой III, отличается от промотора гена U2-РНК, с которым взаимодействует РНК-полимераза II, только отсутствием ТАТА-элемента U2-промотора. У растений оба этих промотора содержат такие регуляторные элементы, а изменение расстояния между ТАТА-элементом и расположенным перед ним регуляторным элементом превращает промотор U6 из Pol III-зависимого в Pol II-зависимый. Несмотря на наличие у промоторов этих РНК-полимераз характерных черт, описанных выше, механизмы, по которым РНК-полимеразы II и III различают свои промоторы, остаются невыясненными.
Промоторы РНК-полимеразы I. Несмотря на то, что участки генома, кодирующие рРНК, относятся к одним из наиболее консервативных, регуляторные последовательности, обеспечивающие инициацию транскрипции у разных видов, сильно дивергированы. Одно из объяснений этого явления основывается на факте вхождения рДНК в мультигенное семейство (гены рРНК многократно повторены в геноме). В этой связи мутации в регуляторной области одного гена, которые благоприятно сказываются на его экспрессии, могут быстро распространиться в семействе последовательностей благодаря генной конверсии, неравному кроссинговеру и/или последовательным актам их эксцизии и реинтеграции в геном. В соответствии с этой интерпретацией, промоторные последовательности рДНК обнаруживают мало гомологии даже у таких близких видов, как человек и мышь. Тем не менее, общий план строения промоторов рДНК практически не различается у всех исследованных эукариот. Промоторы РНК-полимеразы I содержат два кό ровых регуляторных элемента (CPE – core promoter element), локализованных между положениями –75 и –50, а также –30 и +1 (см. рис. I.4, б). Основной промотор лишь слабо связывает TBP-содержащий белковый комплекс SL1 (у людей и крыс), в состав которого, кроме того, входят еще три видоспецифических фактора транскрипции (TBP associated factor – TAF), специфичных в отношении РНК-полимеразы I. Сборка прединициационного комплекса (см. раздел 2.1.3) в этом случае не зависит от специфического взаимодействия TBP и ДНК, так как промоторы РНК-полимеразы I не содержат TATA-последовательности. Как уже упоминалось выше, TAFI63 и TAFI110, входящие в состав комплекса SL1, являются ДНК-распознающими компонентами комплекса. С помощью футпринтинга (см. раздел 7.6.4) было установлено, что комплекс SL1 остается связанным с промотором на протяжении нескольких раундов инициации синтеза РНК РНК-полимеразой I. Стабильное взаимодействие комплекса SL1 с промотором требует участия дополнительного ДНК-связывающего белка UBF, который сам по себе обладает способностью слабо связываться с удаленной регуляторной последовательностью UCE (upstream control element), расположенной между нуклеотидами в положениях –90 и –150. Синергизм в стимулирующем действии SL1 и UBF на транскрипцию РНК-полимеразой I может быть обусловлен наличием непосредственных контактов между этими двумя факторами. Другим регуляторным элементом, удаленным от точки инициации транскрипции промотора Pol I, является энхансер, построенный из коротких повторяющихся последовательностей.
Последовательность CPE обеспечивает специфичность транскрипции генов рРНК, и ее достаточно для инициации. В этой связи последовательность UCE и энхансер не рассматриваются в настоящее время как части промотора РНК-полимеразы I. Кроме того, соответствующие промоторы простейших, грибов и растений последовательность UCE не содержат, а белок UBF не является необходимым компонентом бесклеточных систем транскрипции. Как будет подробно рассмотрено ниже, образование прединициационного комплекса РНК-полимеразой II требует участия фактора TFIIB. Аналогичный в функциональном отношении белковый фактор BRF также необходим для инициации транскрипции РНК-полимеразой III. Соответствующий белковый компонент РНК-полимеразы I еще не идентифицирован. Помимо фактора, связывающего к ó ровую последовательность – CPBF, в инициации синтеза рРНК принимает участие фактор, связывающий энхансер 1, – E1BF (enhancer 1-binding factor), который не входит в состав холофермента Pol I. Оба фактора заметно повышают базальный уровень синтеза рРНК в бесклеточных системах транскрипции.
|
|