Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Спермий Спермий
|
|
Рис 4. Схема развития мужского гаметофита и гамет (на примере сосны).
Пыльцевая трубка, растущая через ткань нуцеллуса по направлению к архегонию, разрушает шейковые канальцевые клетки, конец ее лопается, и спермии попадают к яйцеклетке. Ядро одного из них сливается с ядром яйцеклетки, другой спермий погибает.
Образовавшееся ядро зиготы без периода покоя начинает делиться. Для большинства голосеменных характерна стадия свободноядерного деления, продолжительность которой сильно варьирует у разных групп. У большинства хвойных она очень короткая, клеточные оболочки появляются уже на восьмиядерной стадии – это этап развития «проэмбрио». У зрелого зародыша имеются зародышевый корень, гипокотиль, семядоли (2 и более, до 18), которые окружают точку роста побега.
Зародыш окружен массивной тканью женского гаметофита – первичным эндоспермом, клетки которого содержат крахмал, белки и жиры. Семяпочка превращается в семя. Из интегумента образуется семенная кожура. Остатки нуцеллуса сохраняются только возле микропилярного конца семени. У многих голосеменных при детальном рассмотрении можно обнаружить тонкий внутренний слой покровов семени, который представляет собой некротированную спородерму (мегаспоровую мембрану). Крылатый придаток, который бывает у семян некоторых сосен, елей, кедра, отслаивается от семенной чешуи.
Формирование семени у большинства голосеменных завершается на материнском растении, поэтому отделившееся семя некоторое время остается в состоянии покоя. Однако у многих примитивных вымерших групп семяпочки отделялись от материнского растения еще до момента полного формирования зародыша, а возможно, и до образования зиготы, но с попавшим в пыльцевую камеру мужским гаметофитом. Завершение всех процессов, ведущих к образованию нового спорофита, вероятно, протекало вне материнского растения.
По вопросу классификации голосеменных существуют различные мнения и предлагаются разные варианты. Так, А.Л. Тахтаджян (1986) делит голосеменные на 5 отделов, в составе которых насчитывается 10 классов и 28 порядков.
Система голосеменных по А.Л. Тахтаджяну (1986) (до классов):
Отдел I Lyginopteridophyta (Pteridospermatophyta)
Отдел II. Ginkgophyta
Кл. Ginkgoopsida (2 порядка)
Отдел III. Pinophyta
Кл. 1. Cordaitopsida (1 порядок)
2. Pinopsida (10 порядков)
Отдел IV. Cycadophyta
Кл.1. Cycadopsida (1 порядок)
2. Pentoxylopsida (1 порядок)
3. Cycadeoideopsida ( Bennettitopsida) (1 порядок)
Отдел V. Gnetophyta
Кл. 1. Ephedropsida (1 порядок)
2. Welwitschiopsida (1 порядок)
3. Gnetopsida (1 порядок)
В классификации С.В. Мейена (1987) сохраняется единый отдел Голосеменные (Pinophyta), включающий 3 класса и 19 порядков.
Система голосеменных по С.В. Мейену (1987):
Отдел. Pinophyta
Кл. I. Ginkgoopsida (10 порядков, в том числе Ephedrales)
Кл. II. Pinopsida (Coniferopsida) (3 порядка)
Кл.III. Cycadopsida (6 порядков, в том числе Bennettitales, Welwitschiales и Gnetales).
В данном пособии принята классификация отдела Голосеменные (Pinophyta, Gymnospermae) по А.Л.Тахтаджяну (Жизнь растений, 1978). Отдел включает шесть классов:
1. Pteridospermatopsida, Lyginopteridopsida
2. Bennettitopsida
3. Cycadopsida
4. Ginkgoopsida
5. Pinopsida, Coniferopsida
6. Gnetopsida, Chlamydospermatopsida
Два первых класса известны только по ископаемым остаткам, остальные включают как вымерших, так и ныне живущих представителей. В составе класса Гнетовидные до недавнего времени указывалось только три современных рода и отмечалось отсутствие сведений о вымерших представителях. Однако В.А.Красилов приводит данные о сделанных находках ископаемых гнетовых, представляющие интерес для филогении.
Голосеменные, насчитывающие, по новейшим данным, ~ 900 видов, распространены по всему земному шару и встречаются на всех континентах, кроме Антарктиды. Они играют ведущую средообразующую роль в экосистемах хвойных лесов, занимающих огромные территории на равнинах умеренной зоны северного полушария, а также в лесном поясе горных систем на всех широтах.
В практическом отношении голосеменные, несмотря на относительно малую численность видов, занимают второе место после покрытосеменных. Они поставляют ценнейший материал для строительства и мебельного производства, являются источником различных технических, лекарственных веществ и косметических средств и др. Некоторые виды находят применение как пищевые растения, у которых используются в пищу семена, сердцевина ствола. Многие голосеменные являются декоративными растениями, применяются в посадках вдоль транспортных магистралей в качестве защитных полос, для закрепления песчаного грунта, как противоэрозионные насаждения. Многие реликтовые и редкие виды стали охраняемыми объектами на мировом или региональном уровне. В «Чырвоную кнiгу Рэспублiкi Беларусь» занесена пихта белая (Abies alba).
Класс Семенные папоротники, или Птеридоспермы (Lyginopteridopsida, Pteridospermatopsida)
Семенные папоротники – наиболее древняя вымершая группа голосеменных, существовавшая с позднего девона до раннемелового периода, расцвет ее был в карбоне и перми. Свое название группа получила из-за сочетания признаков папоротников и семенных растений.
Птеридоспермы впервые были описаны в 1904 г. английскими палеоботаниками Ф. Оливером и Д. Скоттом. С папоротниками их сближает сходство по листьям, улиткообразно скрученным в молодом состоянии, позже вильчато раздвоенным и многократно перисторассеченным. Однако наличие семяпочек, а также анатомическое строение стеблей, способных к вторичному утолщению за счет камбия, свидетельствует о принадлежности к голосеменным. Семенные папоротники были распространены по всему земному шару, но отдельные группы имели определенную географическую приуроченность. Например, користоспермовые и глоссоптериевые занимали территорию древнего материка – Гондваны. Семенные папоротники известны из каменноугольных отложений Европы, Азии и Северной Америки.
| Рис. 5. Медуллоза Ноэ (Medullosa noei). 1 – общий вид, реконструкция; 2 – поперечный срез через стебель с тремя меристелами. |
Многие семенные папоротники были древовидными растениями с прямостоячим моно- или дихоподиально ветвящимся стволом до 20 см в диаметре. Листья очередные, сближенные на верхушке (рис. 5). У других представителей стебель был лазающим или лиановидным, 2-4 см в диаметре, с расставленными листьями. Стебли несли придаточные корни, отходившие от узлов и междоузлий. Поверхность стеблей была гладкой или покрыта придатками в виде железистых волосков, основаниями листьев и остатками листовых рубцов. Стебли некоторых групп не известны. Среди семенных папоротников встречаются и формы с цельными листьями. Так, у широко распространенных в Гондване глоссоптериевых листья располагались на тонких стеблях мутовчато или по спирали и имели цельную листовую пластинку линейной, ланцетной, яйцевидной или лопатчатой формы, были черешковые или сидячие, с сетчатым жилкованием, со средней жилкой или без нее.
Анатомическое строение стеблей изучено не у всех групп семенных папоротников, но имеющиеся в палеоботанических работах описания свидетельствуют о его большом разнообразии. По типу стели отмечены протостеличные, сифоностеличные и эустеличные формы, с проводящими пучками концентрического и коллатерального типа. Варьировало также и соотношение различных по мощности участков сердцевины, древесины и коры. Особый интерес представляют репродуктивные структуры этих древнейших представителей семенных растений. Они отличаются чрезвычайным разнообразием по строению и расположению. Микроспорангии располагались поодиночке либо группами, при этом часто срастались в синангии. У более примитивных форм одиночные микроспорангии находились на концах осей, обильно ветвящихся в разных направлениях. У других свободные микроспорангии составляли радиальные ряды на концах дихотомически ветвящихся веточек. У многих имелись синангии, расположенные на спорангиеносных структурах, образующих чашевидные, колоколовидные или другие, более сложные агрегации. Они прикреплялись к рахисам и сегментам листьев перистого или пальчатого типа (рис. 6). Предпыльца без воздушных мешков либо с 1-2 мешками.
Мужской гаметофит, развивавшийся из микроспоры еще внутри микроспорангия, был многоклеточным (до 20-30 клеток). Предполагают, что мужские гаметы были подвижными. Ни гаусториальных, ни пыльцевых трубок в семяпочках с попавшими в них мужскими гаметофитами не обнаружено. Вероятно, подвижные мужские гаметы, оказавшись в пыльцевой камере семяпочки, с помощью жгутиков перемещались к яйцеклеткам. Оплодотворение и развитие зародыша могло происходить после опадения таких семяпочек, поскольку зародышей в них не найдено.
Интерес представляет строение и формирование семяпочек этих первичных семенных растений. Изучение семяпочек самых древних представителей семенных папоротников показало, что их расположение и строение разнообразно и может дать ключ к пониманию путей эволюции этой важнейшей репродуктивной структуры. Основная его часть – нуцеллус (видоизмененный мегаспорангий), имевший различные покровы. Предполагают, что первичный покров – интегумент возник из стерильных осей, окружавших мегаспорангий в виде частично сросшихся 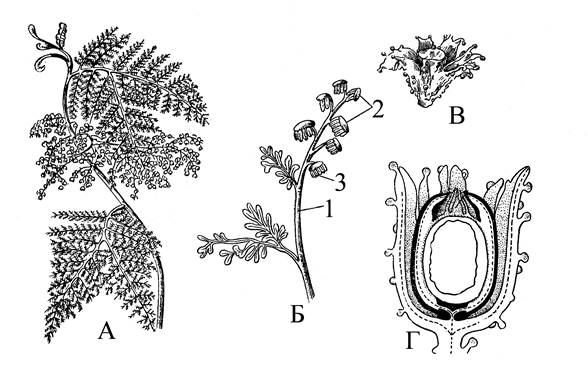 Рис. 6. Калимматотека (Calymmatotheca hoeninghausii): А – общий вид (часть растения); Б – фрагмент листа (1 – вегетативная часть листа; 2 – спорангиеносные сегменты, 3 – микроспорангии); В – семяпочка, окруженная купулой; Г – продольный разрез через семяпочку и купулу.
Рис. 6. Калимматотека (Calymmatotheca hoeninghausii): А – общий вид (часть растения); Б – фрагмент листа (1 – вегетативная часть листа; 2 – спорангиеносные сегменты, 3 – микроспорангии); В – семяпочка, окруженная купулой; Г – продольный разрез через семяпочку и купулу.
лопастей (у некоторых лагеностомовых – Genomosperma). Морфологическая природа этих лопастей, снабженных проводящим пучком, трактовалась по-разному. Ранее их считали стерильными мегаспорангиями (синангиальная гипотеза). В последнее время предполагается возможность их образования из осевых, теломных ветвей. У одних представителей семяпочки не имели дополнительных покровов. У других вокруг семяпочек располагались особые выросты – купулы, многолопастные или чашевидные структуры. Купулы у многих птеридоспермов прирастали к интегументу, образуя внешний интегумент – купуломент. Таким путем возникли двупокровные семяпочки, при этом в каждом покрове сохранялась своя проводящая система. Однако в некоторых случаях внутренний интегумент терял свою проводящую систему и сливался с нуцеллусом, тогда семяпочка становилась вторично однопокровной с единственным, внешним интегументом.
Семяпочки многих семенных папоротников прикреплялись к безлистным осям, образуя простые агрегации кистевидной, колосовидной, головчатой, зонтиковидной, перистовидной и иной формы. У ряда представителей они располагались на листоподобных органах и их сегментах. У некоторых представителей имелись особые полузамкнутые вместилища семяпочек – капсулы (у кейтониевых).
Семяпочки семенных папоротников различаются по типу симметрии: радиально симметричные и билатеральные, или, по терминологии С.В. Мейена, радиоспермические и платиспермические. У более поздних представителей появились вторично-платиспермические формы. Тип симметрии семяпочек используется при классификации голосеменных, в частности, в системе С.В.Мейена как один из ведущих признаков.
В состав класса Семенные папоротники входят: каламопитиевые, лагеностомовые (лигиноптеридиевые), медуллозовые (тригонокарпусовые), пельтаспермовые, глоссоптериевые (арбериевые), кейтониевые и др. В системах разных авторов эти группы имеют разный таксономический ранг и положение. Многие ученые отмечают сборный характер класса и проблематичность его выделения как единого таксона.
Различные группы семенных папоротников сыграли, несомненно, большую роль в эволюции голосеменных и, возможно, покрытосеменных растений. Однако в вопросах конкретных филогенетических связей этого класса очень много неясного, во взглядах ученых имеется немало противоречий. На сегодняшний день нет единого мнения относительно путей эволюции этого таксона.
Класс Саговникоподобные(Cycadopsida)
Саговникоподобные – одна из древнейших групп семенных растений, появившаяся, по-видимому, в пермский период, представители которой дожили до наших дней. Расцвет саговниковых приходится на триасовый и юрский периоды мезозойской эры. В настоящее время от некогда обширной группы осталось всего 10 родов.
По внешнему облику, морфологическим и анатомическим признакам саговниковые выделяются среди современных голосеменных большим своеобразием. Прежде всего, это крупнолистные формы с перистыми (у большинства) и дваждыперистыми (у бовении) листьями с верхушечным перышком, придающими растениям папоротниковидный облик. У бовении молодые листья свернуты улиткообразно, у других они прямые или согнутые (стангерия), с прилистниками (у стангерии прилистник 1). Молодые листья опушены простыми двуклеточными и ветвящимися волосками. Перышки листа различны по форме: линейные, ланцетные, языковидные, ромбовидные, веерные; цельнокрайные или зубчатые. Жилкование параллельное, дихотомическое, иногда вдоль края петлевидное, у некоторых жилки образуют анастомозы.
Для эпидермиса характерно образование чередующихся полос разнонаправленных клеток. Стенки клеток извилистые или прямые. Устьица гаплохейльные, расположены группами, рядами, с почти непогруженными или глубоко погруженными замыкающими клетками. На боковых выростах их иногда заметна радиальная штриховка, как у хвоща. Расположение устьиц – на нижней или на обеих сторонах листа.
Среди саговниковых имеются растения с прямыми неветвящимися коллоновидными стволами (высокорослые – до 10-20 м, низкорослые от 2-3 до 6-9 м), а также формы с клубневидно утолщенными подземными, полунадземными стеблями, несущими на верхушке крупные листья. Растение обычно дает пучок листьев ежегодно или через год, у стангерии отрастает по одному листу в течение года. У обитателей саванн листья сбрасываются в сухое время года. После опадения их на стволах остается чехол из оставшихся оснований. У двух видов рода замия наряду с обычными наземными растениями встречаются отдельные особи, которые растут как эпифиты на деревьях (рис. 7).
У большинства саговниковых имеется удлиненный главный корень, от которого отходят многочисленные боковые, несущие на конце массивный корневой чехлик. В ряде случаев главный корень рано прекращает рост, формируется система придаточных корней, отходящих от основания стебля. У некоторых видов есть контрактильные (втягивающие) корни, которые препятствуют выталкиванию молодого ствола над поверхностью почвы. Своеобразием корневой системы всех саговниковых является наличие особых коралловидно разрастающихся окончаний корней (кораллоиды), которые формируются в результате дихотомического ветвления некоторых боковых корней. В кораллоидах поселяются различные симбионты, в том числе цианеи (Nostoc punctiforme, Anabaena cycadea и др.), азотфиксирующие бактерии (Azotobacter, Bacterium radicicola), некоторые фикомицеты. Это редкий случай многокомпонентного симбиоза у растений. Характер взаимоотношений слагающих его организмов не вполне ясен.
Стебель саговниковых по анатомическому строению относится к маноксильному типу (от греч. manos – рыхлый, неплотный; xylon – древесина). Вторичная древесина, образующаяся за счет деятельности камбия, занимает узкую полосу на поперечном разрезе стебля. Центральная часть его образована сердцевиной (до 1/3 объема).
Протоксилема стебля состоит из трахеид со спиральными утолще-ниями, метаксилема образуют преимущественно лестничные трахеиды, а в зрелой древесине находятся трахеиды с округлыми окаймленными порами без торуса. Характерна значительная длина трахеид (от 7 до 10 мм).

Рис. 7. Формы роста саговниковых.
1 – микроцикас красивокронный (Microcycas calocoma); 2 – энцефаляртос поперечно-жильчатый (Encephalartos transvenosus); 3 – саговник поникающий (Cycas revoluta); 4 – энцефаляртос превосходный (E. princeps); 5 –макрозамия спиральная (Macrozamia spiralis); 6 – замия кремнистая (Zamia silicea); 7 – бовения мелькопильчатая (Bowenia serrulata); 8 – замия паразитная (Z. poeppigiana).
У некоторых представителей наблюдается полициклическое строение стебля. Утолщение стебля при этом происходит за счет последовательного образования в коре добавочных камбиальных колец, которые формируют дополнительные зоны коллатеральных пучков. У саговника образуются также концентрические проводящие пучки в коре. Флоэма представлена ситовидными клетками с рассеянными ситовидными полями и косыми конечными стенками. Имеются также особые сопутствующие клетки и удлиненные волокна.
Массивная сердцевина образована крупными тонкостенными паренхимными клетками, содержащими крахмал. Сердцевина сообщается с корой широкими радиальными сердцевинными лучами. В сердцевине некоторых саговниковых формируется система взаимосвязанных проводящих коллатеральных пучков и связанных с ними слизевых каналов.
Паренхима коры богата крахмалом. В коре находятся слизевые каналы. За счет пробкового камбия образуется толстый слой перидермы.
Все саговниковые являются двудомными растениями. Репродуктивные органы их представлены простыми микростробилами и мегастробилами, только у рода саговник компактный мегастробил не образуется, а имеются мегаспорофиллы, рыхло расположенные на верхушке ствола.
 Микростробилы саговниковых состоят из оси, на которой находятся микропорофиллы, расположенные по спирали или мутовчато. Размеры микростробила у разных представителей сильно различаются: от 2 см длиной (замия флоридская) до 80-90 см (энцефаляртос, макрозамия).
Микростробилы саговниковых состоят из оси, на которой находятся микропорофиллы, расположенные по спирали или мутовчато. Размеры микростробила у разных представителей сильно различаются: от 2 см длиной (замия флоридская) до 80-90 см (энцефаляртос, макрозамия).
| Рис. 8. Репродуктивные структуры саговниковых. Мегаспорофиллы (а – саговника поникающего; б – с. завитого; в – с. Норманбина; г – диоона съедобного); микроспорофиллы (д – саговника завитого; е – замии цельнолистной); ж – группы микроспорангиев замии цельнолистной; з – стенка микроспорангия стангерии (1 – устьица). |
Микроспорофиллы более-менее плоские, чешуевидные или щитовидные (пельтатные) (рис. 8). На нижней поверхности образуются микроспорангии, скученные группами по 2-3-6, частично или полностью срастающиеся в сорусы. Общее число их может достигать 700 и более, но у замии флоридской на отдельных микроспорофиллах может развиваться всего один микроспорангий.
Развитие микроспорангия у саговниковых происходит из группы инициальных клеток (т. е. аналогично эуспорангиатным папоротникам), стенка спорангия многослойная. В эпидермисе микроспорангия у большинства саговниковых есть устьица, что не характерно для голосеменных (кроме саговниковых, это отмечено лишь у некоторых семенных папоротников). Эпидермальный слой пред
ставлен клетками с неравномерно утолщенной клеточной оболочкой, наружная стенка ее остается тонкой. С помощью эпидермальных клеток происходит вскрывание зрелого спорангия продольной трещиной.
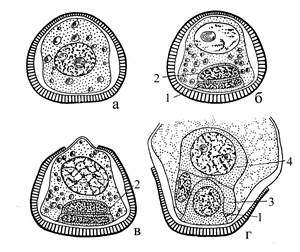
| Рис. 9. Начало развития мужского гаметофита. а – микроспора; б – г – развивающийся гаметофит: 1 – проталлиальная, 2 – антеридиальная, 3 - стерильная, 4 – сперматогенная клетки. |
Из материнских клеток микроспор (микроспороцитов) в результате мейотического деления образуются тетрады микроспор, иногда долго не распадающиеся на одиночные споры. Прорастание микроспоры и формирование мужского гаметофита начинается в спорангии (рис. 9). Вначале образуется одна линзовидная проталлиальная клетка, которая гомологична остатку вегетативной части мужского заростка, и более крупная антеридиальная клетка, обе остаются заключенными в оболочке развивающейся микроспоры. Антеридиальная клетка затем делится на две – генеративную и гаусториальную клетки. В таком 3-клеточном состоянии пыльцевое зерно высыпается из микроспорангия и переносится на семяпочку. Пыльца без воздушных мешков, переносится ветром и, возможно, насекомыми (энцефаляртос).
Мегастробилы саговниковых (женские шишки) по общему плану строения схожи с микростробилами. На общей оси стробила расположены листовидные или щитковидные мегаспорофиллы с семяпочками под щитком. У саговника мегаспорофиллы листовидные, у некоторых видов с перистолопастной, у других с цельной верхушкой, несущие в нижней части по бокам семяпочки (2-8). Они не образуют стробила, а расположены на верхушке стебля. В начале своего развития они образуют подобие шишки, но по мере роста загибаются книзу и свисают вдоль ствола. У видов, имеющих шишковидные мегастробилы, закладка их происходит на верхушке стебля, но в дальнейшем они могут смещаться в пазушное положение, а верхушка продолжает свой рост как симподиальный побег; у 2 родов положение шишек изначально боковое, а рост моноподиальный. У многих саговниковых мегастробилы крупные, до 1 м в длину и до 40-45 кг (энцефаляртос кафрский, лепидозамия Перовского), однако есть и очень мелкие – 2-5 см в длину (замия карликовая).
Семяпочки саговниковых различны по размерам: от 0, 5-0, 7 см (у замии карликовой) до 5-6 см (у энцефаляртоса), однако по строению они однотипны. Основную часть молодой семяпочки составляет нуцеллус, его окружает многослойный интегумент, в котором выделяется внутренний, постепенно рассасывающийся мясистый слой, средний, каменистый слой (склеротеста) и сочный наружный слой (саркотеста), покрытый кожурой. В эпидермисе интегумента имеются устьица. Часто на верхушке интегумента располагаются волоски. Интегумент снабжен двойной проводящей системой, по восемь пучков во внешнем и внутреннем слое. Внутренние пучки могут заходить и в нуцеллус, который плотно прилегает к интегументу на всем протяжении, кроме верхушки, где формируется пыльцевая камера под микропиле. На верхушке нуцеллуса имеется слой кутикулы ~ 7 мкм и очень тонкий слой кутикулы вдоль стенки, прилегающей к интегументу.
По мере формирования семяпочки в нуцеллусе происходят процессы, связанные с образованием мегаспор. Одна из клеток нуцеллуса увеличивается в размерах и становится материнской клеткой мегаспор (мегаспороцит). В результате мейоза формируется линейная тетрада мегаспор. Функциональной является только нижняя мегаспора, а три другие дегенерируют. В результате многократных делений ядра самой мегаспоры и его производных образуется вначале сводобноядерный, а затем клеточный женский гаметофит – первичный эндосперм, заполняющий внутреннюю часть семяпочки. В верхней, обращенной к микропиле части гаметофита формируются архегонии, число которых варьирует от 2-4, реже 3-5 (до10). У микроцикаса образуется до 200 архегониев, расположенных по всей периферии женского гаметофита. Однако в дальнейшем функционирует обычно только один гаметангий. Архегоний саговниковых сильно редуцирован и состоит из яйцеклетки и двух шейковых клеток, которые слегка выдаются в архегониальную камеру, расположенную над архегониями. Ядро брюшной канальцевой клетки быстро дегенерирует. Непосредственно перед оплодотворением число шейковых клеток удваивается. Следует отметить, что в клетках эндосперма имеются лейкопласты, которые могут превращаться в хлоропласты, вызывая «позеленение» ткани заростка, например, в семяпочках, где не произошло оплодотворение яйцеклетки. Это можно оценить как примитивный признак. К числу таковых признаков относится и наличие довольно массивной (до 10 мкм), долго сохраняющейся мегаспоровой оболочки вокруг женского гаметофита в семяпочке саговниковых. Это так называемая мегаспоровая мембрана, сильно трансформированная по сравнению с ее первоначальным состоянием в виде оболочки мегаспоры. На микропилярном полюсе в результате разрушения мегаспоровой мембраны происходит объединение архегониальной и пыльцевой полости в общую пыльцевую камеру, куда попадает пыльцевое зерно из микропиле (рис. 10).
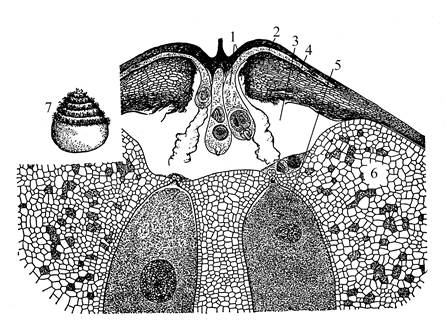 Пыльцевое зерно, попавшее в пыльцевую камеру семяпочки, начинает прорастать. Гаусториальная клетка вытягивается в виде длинного гаустория, который внедряется в ткань нуцеллуса и доставляет питательные веще-
Пыльцевое зерно, попавшее в пыльцевую камеру семяпочки, начинает прорастать. Гаусториальная клетка вытягивается в виде длинного гаустория, который внедряется в ткань нуцеллуса и доставляет питательные веще-
| Рис. 10. Верхняя часть семяпочки диоона (Dioon edule) с прорастающими пыльцевыми зернами. 1 – проросшее пыльцевое зерно; 2 – гаусториальная трубка; 3 – пыльцевая камера; 4 – интегумент; 5 – архегоний; 6 – эндосперм; 7 – сперматозоид. |
ства клеткам развивающегося мужского гаметофита. Из генеративной клетки в результате митотического деления образуются «клетка-ножка» (стерильная, или сестринская) и сперматогенная клетки.
Сперматогенная клетка делится и дает 2 сперматозоида. Отмечено, что у микроцикаса иногда делится также и «клетка-ножка», дающая линейный ряд сперматогенных клеток, но, чаще всего, она остается стерильной.
Сперматозоиды саговниковых крупные, подвижные, несущие на особом придатке-блефаропласте огромное количество жгутиков (до 20 тыс.). Сперматозоиды были впервые открыты и описаны у саговника развернутого японским ботаником С. Икено в 1896 – 1898 гг. и независимо от него в 1897–1901 гг.– у замии Веббером.
Между опылением и оплодотворением у разных представителей саговниковых проходит от 4 (у саговника) до 6 (у диоон) месяцев. Перед оплодотворением сперматозоиды вместе с небольшим количеством жидкости перемещаются в архегониальную камеру семяпочки. При помощи амебоидных и пульсирующих движений происходит их вхождение в яйцеклетку. Большая часть сперматозоида вместе с блефаропластом остается за пределами архегония. Функционирует только один из двух сперматозоидов. После слияния яйцеклетки и сперматозоида образовавшаяся зигота без периода покоя начинает делиться. Однако процессы, связанные с развитием зародыша, происходят обычно после опадения семяпочки с материнского растения.
Первые этапы эмбрионального развития характеризуются делением ядер без образования клеточных перегородок. У диоона число свободных ядер достигает ~ 1000. В дальнейшем появляются перегородки, формируется многоклеточный предзародыш. В нижней части его образуется спирально скрученный подвесок (суспензор), а на верхнем конце развивается зародыш, который по мере разрастания продвигается подвеском в ткани эндосперма. В зародыше формируется зародышевый корешок, окруженный корневым влагалищем – колеоризой, обращенной к микропилярному концу. За ним следует подсемядольное колено – гипокотиль с двумя (саговник) или несколькими семядолями, между которыми находится почечка. У диоона 3-4 семядоли, у микроцикаса 3-6, а у цератозамии только 1, вторая дегенерирует.
В процессе формирования семени в клетках эндосперма накапливаются питательные вещества – крахмал, масла, белки за счет нуцеллуса и внутреннего слоя интегумента, которые постепенно рассасываются. Наружный слой интегумента образует сочную часть семени, покрытую кожурой, средний каменистый дает твердую «косточку» (наподобие костянки сливы).
При прорастании зрелого семени первой наружу «пробивается» колеориза, сквозь нее прорастает корень. Семядоли не выносятся на поверхность (подземное прорастание), они доставляют питательные вещества из эндосперма развивающемуся проростку. Рост молодого растения происходит обычно медленно, только спустя 5-6 лет формируется первая мутовка листьев.
В современной флоре класс Саговникоподобные (Cycadopsida) представлен одним порядком Саговниковидные (Cycadales), в составе которого иногда выделяют 3 семейства – Саговниковые (Cycadaceae), Стангериевые (Stangeriaceae) и Замиевые (Zamiaceae). Некоторые ботаники все 10 родов, включающих 120-130 видов, рассматривают как одно семейство Саговниковые – Cycadaceae.
Саговниковые обитают в тропических и субтропических областях обоих полушарий. Выделяются три обширные зоны родового эндемизма: американская, где распространены роды замия (Zamia), цератозамия (Ceratozomia), диоон (Dioon) и микроцикас (Microcycas); африканская – с родами энцефаляртос (Encephalartos), стангерия (Stangeria) и наиболее обширная азиатско-австралийская зона, в которой представлены саговник (Cycas), основной ареал которого азиатский, но заходит в Африку и на Мадагаскар, и три австралийских рода – макрозамия (Macrozamia), лепидозамия (Lepidozamia) и бовения (Bowenia). Большинство видов саговниковых произрастает на приокеанских территориях материков и островов, лишь немногие виды проникают вглубь материка. Саговниковые являются обитателями низкорослых вечнозеленых лесов и кустарниковых зарослей, другие встречаются в дождевых тропических лесах (лепидозамия Хоупта, некоторые замии в лесах вдоль Амазонки), на открытых местах в саваннах Африки и Америки, на обрывах и скалах морских побережий (саговник поникающий), на коралловых рифах (замия флоридская). Два вида замий (Zamia poeppigiana и Z. pseudoparasitica) являются факультативными эпифитами, поселяясь как на почве, так и на стволах и ветвях деревьев, не будучи, однако, паразитами. Стебель их высотой до 1, 5 м, зеленые листья до 2 м в длину.
Практическое значение имеют некоторые виды саговниковых, используемые как пищевые, лекарственные, пряноароматические, декоративные и др. Из сердцевины энцефаляртоса получают крупу саго, из эндосперма семян – крахмал, масло, сочные листья употребляют как овощ или в качестве пряной добавки (керри-пряность). Некоторые виды выращивают как декоративные в оранжереях и открытом грунте, листья саговника используют для ритуальных целей (на погребальные венки). В связи с узким эндемизмом некоторых видов, сокращением численности многих возникла угроза исчезновения, поэтому необходима охрана этих оригинальных и уникальных представителей относительно примитивной группы голосеменных.
В проблеме происхождения класса Cycadopsida много неясного. Ископаемые остатки голосеменных, определяемые как принадлежащие к цикадофитам, оказываются близкими к разным группам семенных папоротников (лигиноптериды, медуллозовые). Принадлежность к саговниковым многих описанных палеоботаниками форм после уточнения подвергается сомнению или вовсе отвергается. По-разному трактуются направления эволюции, морфологическая природа различных структур, в первую очередь репродуктивных. Поэтому вопрос о предках этого класса остается нерешенным. По мнению В.А.Красилова (1989), среди гетерогенного таксона семенные папоротники существовала группа Protocycadopsida, которая явилась родоначальной для более поздних саговниковых. Современные роды этого класса появились не раньше юрского и мелового периодов.
Класс Беннеттитоподобные (Bennettitopsida)
 Беннеттитовые являются более поздней, чем семенные папоротники, группой голосеменных растений, занимавшей господствующее положение от позднего триаса до середины мелового периода и вымершей к его концу.
Беннеттитовые являются более поздней, чем семенные папоротники, группой голосеменных растений, занимавшей господствующее положение от позднего триаса до середины мелового периода и вымершей к его концу.
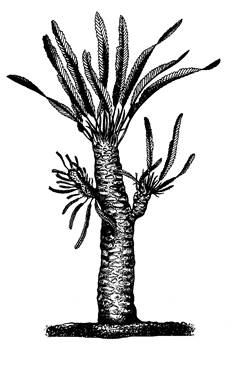
| Рис.11. Внешний вид Вильямсонии Сьюорда (Williamsonia sewardiana). Реконструкция. |
Беннеттитовые были древовидными растениями разнообразного внешнего облика. Одни могли достигать размеров высокого дерева с верхушечной кроной, другие были кустарниковыми формами либо имели невысокие цилиндрические или клубневидно утолщенные неветвящиеся стволики высотой от нескольких сантиметров до 3-4 м. Различают лептокаульную (тонкоствольную) и пахикаульную (толстоствольную) формы роста. К первому типу относится род виеландиелла (Wielandiella) – кустарниковая форма, имевшая стебли около 2 см толщиной, которые симподиально ветвились, образуя боковые побеги 0, 5-1 см в диаметре.
К лептокаульным относятся виллиямсония (Williamsonia) (рис. 11) и виллиямсониелла (Williamsoniella) (рис. 12), имевшие довольно высокие почти не ветвящиеся утолщенные стволы (описаны под названием Bucklandia) покрытые неопадающими листовыми черешками либо остатками опавших листьев. Возможно, у некоторых беннеттитовых происходил сезонный листопад. Остатки таких беннеттитовых обнаружены в различных местах обитания: в карбонатных осадках лагун и соленых озер, на пресноводных болотах и в кустарниковых зарослях, где встречались также хвойные и гнетовые (Красилов, 1989). Пахикаульные формы росли, очевидно, на сухих открытых местах.
| Рис. 12. Внешний вид Williamsoniella coronata (реконструкция). а – внешний вид; б – обоеполый стробил; в - микроспорофилл. |
 Пахикаульная жизненная форма описана на примере цикадеоидеи (Cycadeoidea). Невысокие клубневидные стволики росли одиночно или группами по нескольку, были тесно прижаты друг к другу, возможно, они развивались из почек одного подземного побега. Поверхность такого стебля была покрыта пластинчатыми чешуйками или волосками, а также остающимися черешками опавших листьев. Перистые листья располагались на верхушке в виде пучка (рис. 13).
Пахикаульная жизненная форма описана на примере цикадеоидеи (Cycadeoidea). Невысокие клубневидные стволики росли одиночно или группами по нескольку, были тесно прижаты друг к другу, возможно, они развивались из почек одного подземного побега. Поверхность такого стебля была покрыта пластинчатыми чешуйками или волосками, а также остающимися черешками опавших листьев. Перистые листья располагались на верхушке в виде пучка (рис. 13).
 Анатомическое строение стеблей характеризуется наличием эустели с эндархной протоксилемой из спиральных и с метаксилемой из лестничных трахеид. Вторичная ксилема состояла из лестничных и (или) точечных трахеид с окаймленными порами. Отмечены также узкие сердцевинные лучи. В коре находились мелкие мезархные пучки, заходившие в листья, центральную часть стебля занимала обильная
Анатомическое строение стеблей характеризуется наличием эустели с эндархной протоксилемой из спиральных и с метаксилемой из лестничных трахеид. Вторичная ксилема состояла из лестничных и (или) точечных трахеид с окаймленными порами. Отмечены также узкие сердцевинные лучи. В коре находились мелкие мезархные пучки, заходившие в листья, центральную часть стебля занимала обильная
| Рис. 13. Внешний вид Cicadeoidea marshiana (реконструкция). |
сердцевина.
Листья беннеттитовых двух типов – простые с цельной или рассеченной листовой пластинкой и сложные перистые с линейными или продолговатыми боковыми листочками, пластинки которых расположены наклонно к плоскости листа, как жалюзи. Листья часто обильно опушены многоклеточными и звездчатыми волосками. Сложные листья были похожи на листья саговниковых, что нередко приводило к путанице при определении ископаемых остатков. Однако схожие по внешнему виду листья этих двух классов хорошо распознаются по строению устьичного аппарата и эпидермальных клеток листа. Устьица беннеттитовых синдетохейльные, их замыкающие и побочные клетки развиваются из одной зачаточной клетки. Наружные стенки замыкающих клеток кутинизированы. У саговниковых устьица гаплохейльные, их замыкающие и побочные клетки образуются из разных инициальных клеток. Замыкающие клетки с одревесневшими оболочками (лигнифицированы). Кроме того, клетки эпидермиса листа у беннеттитовых с извилистыми складчатыми стенками, а у саговниковых стенки эпидермальных клеток прямые.
Реконструкция общего облика сделана только для отдельных представителей беннеттитовых, для большинства дается описание отдельных органов и их частей. Особый интерес представляют репродуктивные структуры беннеттитовых, поскольку это единственная группа среди голосеменных, имевшая в своем составе представителей с обоеполыми стробилами. Однако следует иметь в виду, что многие беннеттитовые имели однополые стробилы и были, вероятно, как однодомными, так и двудомными растениями. Стробилы располагались в большом числе в пазухах листовых черешков вдоль утолщенного клубневидного ствола преимущественно в верхней части. У кустарниковых форм стробилы прикреплялись к верхушке центральной оси дихазия.
Наибольший интерес представляют обоеполые стробилы, обнаруженные палеоботаниками в конце прошлого века. Открытие обоеполых стробилов у беннеттитовых дало основание для разработки стробилярной гипотезы происхождения цветка.
Предполагается, что на удлиненной оси обоеполого стробила внизу по спирали располагались опушенные цельные или перистые листья, выше находились мутовки микроспорофиллов со свободными или сросшимися в синангии микроспорангиями. Конусовидная верхушечная часть оси стробила несла мегаспорофиллы с одной семяпочкой на конце, чередующиеся с бесплодными межсеменными чешуями.
По реконструкции стробила Cycadeoidea dacotensis, сделанной Виландом (Wieland), перистые микроспорофиллы изображены в улиткообразно свернутом и развернутом виде. Уточненные модели стробила показывают, что в расправленном состоянии микроспорофиллы не найдены, они всегда согнуты. В нижней части микроспорофилл нес отростки, к которым прикреплялись 2 ряда синангиев из удлиненных спорангиев. Зрелые синангии раскрывались двумя створками. Известны также мегаспорофиллы, расположенные в мутовке и чашевидно сросшиеся между собой. Семяпочки беннеттитовых одиночные с длинной микропиллярной трубкой, на удлиненных нитевидных ножках или почти сидячие. Они были защищены расширенными верхушками сомкнутых межсеменных чешуй, над которыми выступали концы микропилярных трубок. Интегумент семяпочек сросшийся с нуцеллусом. Иногда вокруг семяпочки имелась свободная купула. У некоторых представителей над семяпочками была корона, образованная расширенной верхушечной частью оси стробила с приросшими к ней рудиментами чешуй или микроспорофиллов. Найдены семена с хорошо сохранившимся зародышем, у которого видны две мясистые семядоли и зародышевый корешок, обращенный к микропилярному полюсу. Эндосперм отсутствовал, т.е. семена «безбелковые», что может рассматриваться как признак эволюционной продвинутости. По этому признаку многие покрытосеменные более примитивны, чем беннеттитовые, с которыми их сближают. Согласно стробилярной гипотезе беннеттитовые предполагались в качестве вероятной предковой группы для цветковых.
В составе класса Bennettitopsida указывается один порядок Bennettitales с тремя семействами. Различия между семействами состоят преимущественно в особенностях жизненных форм представителей.
Предполагается, что в своем происхождении беннеттитовые связаны с семенными папоротниками, из которых к ним ближе других групп медуллозовые (тригонокарповые) и более древние каламопитиевые (гидраспермовые). Сами же беннеттитовые, вероятно, близки к предкам отдела Покрытосеменные, а, возможно, являются прямыми предками только некоторых групп покрытосеменных.
Класс Гинкгоподобные (Ginkgoopsida)
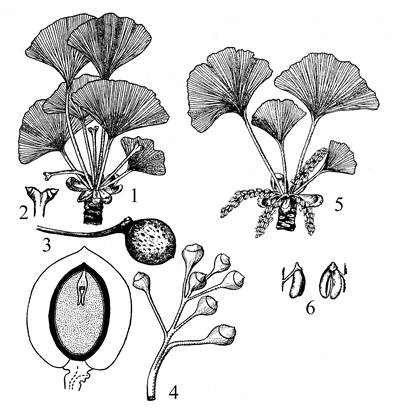
В современной флоре этот класс представлен единственным видом – гинкго двулопастный (Ginkgo biloba), известным в ископаемом состоянии с верхнего триаса. В естественных условиях этот вид сохранился, повидимому, только в Восточном Китае, где на небольшой территории в горах он входит в состав лесной формации вместе с хвойными и широколиственными породами. Издавна известен как священное дерево, культивируемое в Китае, Японии и Корее. В Европу завезен в первой половине 18 века, а в конце столетия – в Северную Америку.
Гинкго двулопастный – листопадное моноподиально ветвящееся дерево высотой до 30 метров. Имеются побеги двух типов: удлиненные – ауксибласты и укороченные боковые – брахибласты. На тех и других располагаются вееровидные листья: с двулопастной пластинкой на удлиненных и почти цельной – на укороченных побегах, где они скучены по 3-5 в пучки. Жилкование дихотомическое, из черешка в листовую пластинку входит два коллатеральных проводящих пучка, которые затем разветвляются, не образуя анастомозов. Устьица гаплохейльные. располагаются на нижней стороне листа, но на листьях удлиненных побегов мужских экземпляров и молодых растений бывают и на верхней стороне.
Ствол и удлиненные побеги пикноксильные, т.е. со слабо развитой сердцевиной и тонкой корой, но мощной вторичной ксилемой из трахеид с окаймленными порами. Древесинная паренхима отсутствует, а сердцевинные лучи шириной в одну клетку. Хорошо выражены годичные кольца. Укороченные побеги маноксильные, сердцевина и кора в них выражены лучше, а древесина мягче, чем в удлиненных побегах.
| Рис. 14. Гинкго двулопастный. 1 – ветвь с мегастробилами; 2 – отдельный мегастробил; 3 – семя; 4 – аномальный ветвистый мегастробил; 5 – ветвь с микростробилами; 6 – микроспорофиллы с микроспорангиями; 7 – семя в разрезе. |
В сердцевине и первичной коре стебля, а также в корне, черешках, листовых пластинках, в семяножке и в семени находятся лизигенные смоляные вместилища, но они отсутствуют во вторичной коре.
 Гинкго двулопастный является двудомным растением, мега- и микростробилы расположены на разных особях. Сильно редуцированные мегастробилы образуются на верхушке укороченных побегов вперемежку с листьями. Короткая ось мегастробила несет на верхушке две супротивно расположенные семяпочки, каждая из них окружена в основании чашевидным выростом неясной морфологической природы. Иногда одна семяпочка бывает недоразвитой, реже ось разветвляется и образуется несколько семяпочек (рис. 14). В основание каждой семяпочки входит пара проводящих пучков. Семяпочка гинкго билатерально симметричная (платиспермическая), с одним интегументом. Наружный слой интегумента сочный, мясистый (саркотеста), в нем проходят два киля в главной плоскости симметрии. Под саркотестой расположен плотный каменистый слой (склеротеста), в котором проходят два проводящих пучка под килями саркотесты. Внутренний слой интегумента тонкий, пергаментный, плотно прилегает к нуцеллусу. В верхней части семяпочки нуцеллус не срастается с интегументом, вследствие чего под микропиле образуется пыльцевая камера. В нуцеллусе происходит мегаспорогенез; из одной материнской клетки мегаспор в результате мейотического деления образуется линейная тетрада мегаспор. Три из них дегенерируют. Четвертая, обращенная к халазальному полюсу мегаспора дает начало женскому гаметофиту. При его развитии в начале отмечается свободноядерная стадия (до 256 ядер), затем в центростремительном направлении идет формирование клеточной структуры гаметофита. На микропилярном полюсе образуется обычно два (реже 3-4) архегония, над которыми находится архегониальная камера. Она отделена от пыльцевой камеры, расположенной выше, тонкой оболочкой мегаспоры, при разрушении которой возникает общая пыльцевая камера под микропиле. В каждом архегонии имеются 4 шейковые канальцевые клетки, которые вдаются в пыльцевую камеру, брюшная канальцевая клетка и яйцеклетка. Остальную часть женского гаметофита составляет обильный эндосперм. Микростробилы, напоминающие по внешнему виду сережчатые соцветия, образуются на верхушке в пазухе ложковидных чешуй укороченных побегов мужских экземпляров. На оси микростробила располагаются по спирали спорангиофоры (спорофиллы), состоящие из короткой ножки и двулопастного щитка, несущего по 2 (реже 3-4) микроспорангия. Стенка микроспорангия многослойная (4-7 слоев). Наружный слой клеток тонкостенный, под ним находится слой клеток с полосатыми утолщениями оболочек – эндотеций, который способствует разрыву стенки продольной трещиной. В микроспорангии образуются эллипсоидальные микроспоры 27-32 мкм в длину, пыльцевые зерна без воздушных мешков.
Гинкго двулопастный является двудомным растением, мега- и микростробилы расположены на разных особях. Сильно редуцированные мегастробилы образуются на верхушке укороченных побегов вперемежку с листьями. Короткая ось мегастробила несет на верхушке две супротивно расположенные семяпочки, каждая из них окружена в основании чашевидным выростом неясной морфологической природы. Иногда одна семяпочка бывает недоразвитой, реже ось разветвляется и образуется несколько семяпочек (рис. 14). В основание каждой семяпочки входит пара проводящих пучков. Семяпочка гинкго билатерально симметричная (платиспермическая), с одним интегументом. Наружный слой интегумента сочный, мясистый (саркотеста), в нем проходят два киля в главной плоскости симметрии. Под саркотестой расположен плотный каменистый слой (склеротеста), в котором проходят два проводящих пучка под килями саркотесты. Внутренний слой интегумента тонкий, пергаментный, плотно прилегает к нуцеллусу. В верхней части семяпочки нуцеллус не срастается с интегументом, вследствие чего под микропиле образуется пыльцевая камера. В нуцеллусе происходит мегаспорогенез; из одной материнской клетки мегаспор в результате мейотического деления образуется линейная тетрада мегаспор. Три из них дегенерируют. Четвертая, обращенная к халазальному полюсу мегаспора дает начало женскому гаметофиту. При его развитии в начале отмечается свободноядерная стадия (до 256 ядер), затем в центростремительном направлении идет формирование клеточной структуры гаметофита. На микропилярном полюсе образуется обычно два (реже 3-4) архегония, над которыми находится архегониальная камера. Она отделена от пыльцевой камеры, расположенной выше, тонкой оболочкой мегаспоры, при разрушении которой возникает общая пыльцевая камера под микропиле. В каждом архегонии имеются 4 шейковые канальцевые клетки, которые вдаются в пыльцевую камеру, брюшная канальцевая клетка и яйцеклетка. Остальную часть женского гаметофита составляет обильный эндосперм. Микростробилы, напоминающие по внешнему виду сережчатые соцветия, образуются на верхушке в пазухе ложковидных чешуй укороченных побегов мужских экземпляров. На оси микростробила располагаются по спирали спорангиофоры (спорофиллы), состоящие из короткой ножки и двулопастного щитка, несущего по 2 (реже 3-4) микроспорангия. Стенка микроспорангия многослойная (4-7 слоев). Наружный слой клеток тонкостенный, под ним находится слой клеток с полосатыми утолщениями оболочек – эндотеций, который способствует разрыву стенки продольной трещиной. В микроспорангии образуются эллипсоидальные микроспоры 27-32 мкм в длину, пыльцевые зерна без воздушных мешков.
Развитие мужского гаметофита из микроспоры начинается в микроспорангии и заканчивается после переноса пыльцы и попадания ее в пыльцевую камеру семяпочки. Процесс формирования мужского гаметофита у гинкго проходит по той же схеме, что и у сосны (рис. 4). Принципиальные отличия заключаются в том, что, во-первых, сифоногенная клетка дает начало гаусториальной, а не пыльцевой трубке, а во вторых, из сперматогенной клетки образуются два подвижных сперматозоида (а не лишенные жгутиков спермии), для которых гаусториальная трубка не является проводником. В четырехклеточном состоянии пыльца переносится на микропиле семяпочки. Благодаря выступающей наружу опылительной капле пыльца удерживается на микропиле и по мере подсыхания жидкости «засасывается» в пыльцевую камеру, где и прорастает. При этом гаусториальная клетка вытягивается в гаусториальную трубку, которая внедряется в ткань нуцеллуса, прикрепляет мужской гаметофит и поставляет питательные вещества образующимся гаметам. Из генеративной клетки образуются сперматогенная и сестринская клетки.
Сперматогенная клетка делится митотически с образованием двух сперматозоидов, несущих многочисленные жгутики на блефаропласте, который находится на одном из полюсов гаметы. Сперматогенез у гинкго впервые подробно проследил и описал Хиразе (Hirase, 1895).
Процесс оплодотворения происходит следующим образом. Сперматозоиды попадают в архегониальную камеру, посредством пульсирующих и амебоидных движений происходит их вхождение в яйцеклетку. Этому способствует образование своеобразного «клюва» за счет выталкивания части цитоплазмы яйцеклетки в зону дегенерирующей брюшной канальцевой клетки. «Клюв» раздвигает шейковые клетки, образуется канал, заполненный жидкостью. Ядро яйцеклетки удлиняется и втягивается в «клюв». При вхождении головки сперматозоида в архегоний и прохождении его через шейковые канальцевые клетки большая часть его тела вместе со жгутиками остается за пределами архегония. При соприкосновении сперматозоида с яйцеклеткой ядро ее перемещается к центру, а «клюв» втягивается внутрь, образуется рецептивная полость, куда попадает головка сперматозоида. Два ядра – мужское и женское – приближаются друг к другу и сливаются, нехроматиновая часть головки сперматозоида полностью абсорбируется цитоплазмой яйцеклетки, образуется зигота.
Обычно в пыльцевой камере семяпочки прорастает несколько пыльцевых зерен, но как только в одной из гаусториальных трубок происходит высвобождение сперматозоидов, другие трубки прекращают свой рост и дегенерируют. При вхождении одного из сперматозоидов в одну из яйцеклеток второй может проникнуть в другую яйцеклетку, однако нормального оплодотворения там не происходит.
При развитии зародыша первоначально бывает стадия свободноядерных делений (до 128–256 ядер). Затем формируется подвесок и зародыш.
Развитие зародыша начинается на материнском растении, но завершается обычно после опадения развивающегося семени с материнского растения.
У зрелого зародыша гинкго две семядоли, которые при прорастании семени не выносятся на поверхность (подземное прорастание). Первые листья проростка чешуевидные, затем появляются нормальные веерные листья.
Осенью наружные сочные покровы семени приобретают янтарно-желтую окраску. Они содержат прогоркающие масла, имеющие неприятный запах. Очищенные от мякоти «косточки» имеют гладкую поверхность, их называют «серебряным абрикосом» (в Японии) и употребляют в пищу их содержимое.
Гинкго двулопастный широко распространен в культуре субтропических и южных районов умеренной зоны в качестве декоративного растения. В ботаническом саду в Киеве растения дают семена. В некоторых парках Беларуси встречаются единичные экземпляры этого экзотического дерева. В настоящее время гинкго используется как лекарственное растение и для получения косметических средств.
Кроме гинкго двулопастного, описан целый ряд ископаемых представителей, наиболее древние – из палеозоя (Sphenobaiera – из перми). Однако многие роды, установленные только по остаткам стеблей и листьев, были отнесены к гинкговым ошибочно. Систематическое положение многих остается неясным.
Классификация этой группы очень противоречивая и нестабильная. Таксономический ранг и объем гинкговых неодинаковы даже в разных вариантах системы одного автора. В принятом здесь варианте системы А.Л.Тахтаджяна (1978) гинкговые принимаются в ранге класса Ginkgoоpsida с одним порядком Ginkgoales.
Класс сосноподобные, или Хвойные, Шишконосные (Pinopsida, Coniferopsida)
Хвойные известны с каменноугольного периода палеозойской эры, расцвет их приходится на юрский и меловой периоды мезозойской эры. К настоящему времени многие группы хвойных полностью вымерли или значительно уменьшились по численности, которая составляет около 630 видов в современной флоре. Они входят в состав 68-ми родов и 8-ми семейств.
Класс хвойные является самой многочисленной группой голосеменных, которая занимает второе место после покрытосеменных по роли в образовании растительного покрова Земли и по значению для человека. Они образуют леса на огромных пространствах в умеренной зоне Северного полушария. Большое видовое разнообразие отмечается в Северной Америке вдоль побережья Тихого океана, где сохранилось много реликтовых видов хвойных. Многие виды встречаются в Юго-Восточном и Центральном Китае, Тайване, Японии, Новой Каледонии, Тасмании, на островах Новой Гвинеи. Ареал 200 видов хвойных ограничен южным полушарием, однако значительные территории, занятуе хвойными лесами, здесь отсутствуют.
Хвойные характеризуются следующими особенностями:
а) древесные жизненные формы, у которых стебли с мощной вторичной древесиной, слабо выраженной сердцевиной и относительно тонкой корой; у хвойных умеренной зоны вторичная древесина образует годичные кольца;
б) стержневая корневая система с многочисленными разветвленными боковыми корнями;
в) наличие смоляных вместилищ в стебле, реже в листьях;
г) мелколистные формы с простыми игловидными, чешуевидными листьями с одной или двумя жилками, реже листья линейные или ланцетные со многими жилками; большинство – с неодновременно опадающими листьями (вечнозеленые растения);
д) микростробилы чаще всего в виде простых шишек (мужские шишки), реже они образуют сложные стробилы;
е) неподвижные (безжгутиковые) мужские гаметы – спермии (у современных представителей), доставка их к яйцеклетке осуществляется с помощью пыльцевой трубки;
ж) мегастробилы образуют сложную шишку, или констробил (женская шишка), реже образуются сильно упрощенные одиночные мегастробилы;
з) семена после созревания на материнском растении и опадения имеют период покоя.
Класс Хвойные (Pinopsida) включает два подкласса:
п/кл. Кордаитиды – (Cordaitanthidae)
п/кл. Пиниды, Хвойные – (Pinidae).
Первый подкласс – полностью вымершая группа, второй представлен как современными, так и ископаемыми таксонами.
Подкласс Кордаитиды – (Cordaitanthidae)
В составе этого подкласса только один порядок Кордаитовые (Cordaitanthales). Геологическая история этой группы берет свое начало с каменноугольного периода. По крайней мере, в конце карбона, т. е. ~ 290 млн. лет назад, кордаиты были широко распространены как в болотистых, так и в более сухих местообитаниях, образуя леса. Они в изобилии были представлены также и на протяжении более сухого и холодного пермского периода.
По своему облику это были деревья до 30 м в высоту с широкими лентовидными листьями до 1 м в длину и от 1 до 15 см в ширину, с тяжами гиподермальной механической ткани. Многие гигрофильные кордаиты представляли собой невысокие кустарники со стелющимся стеблем толщиной около 5 см, с многочисленными придаточными корнями, с укороченными и удлиненными побегами, несущими листья игловидные и лентовидные. Сердцевина стебля у некоторых представителей была разделена вертикальными трещинами (септированная). Проводящая система стебля эустеличная. В листе находятся проводящие коллатеральные пучки в числе от 2 до 8-10. В ксилеме стебля наблюдается постепенный переход от кольчатых, спиральных и лестничных трахеид к точечным с окаймленными порами, главным образом, на радиальных стенках.
У более древних представителей годичные кольца древесины отсутствовали, у более поздних пермских представителей они хорошо выражены. Снаружи стебля находилась относительно тонкая кора с секреторными, вероятно смоляными вместилищами.
Листья были сидячими, линейными или узколанцетными длиной от 10см до 1 м, суженными к основанию и верхушке, тупыми или заостренными. Устьица тетрацитного типа располагались либо только на нижней стороне листа, либо на верхней и нижней в продольных желобках. Тяжи склеренхимы находились между пучками проводящей ткани, образуя «ложные жилки».
Листья кордаитовых, по мнению палеоботаника С.В.Мейена, возникли в результате уплощения черешка (филлодизации) листа, который имелся у листьев папоротникового типа предполагаемых предков кордаитов – лагеностомовых. Перышки листа могли претерпеть полную редукцию.
Репродуктивные органы кордаитовых довольно разнообразны по расположению и деталям строения, но общий план сходный.
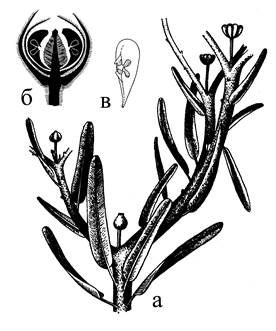
Сложные микростробилы кордаитантуса находились на олиственных побегах вперемежку с листьями. На четырехгранной оси стробила располагались кроющие чешуи – брактеи, образуя 4 ряда. В пазухе каждой брактеи был радиально симметричный укороченный побег, который представлял собой элементарный (простой) микростробил с микроспорофиллами, имевшими по 4 (3-6) микроспорангия на верхушке (рис. 15). Микроспорангии вскрывались продольной трещиной. Пыльцевые зерна с воздушным мешком.
Мегастробилы, изученные более детально у кордаитантуса, располагались на укороченных пазушных побегах. Они несли стерильные чешуи у основания оси и спорофиллы с одной-двумя семяпочками на конце. Простые стробилы собраны в общее собрание – сложный стробил (рис. 18а).
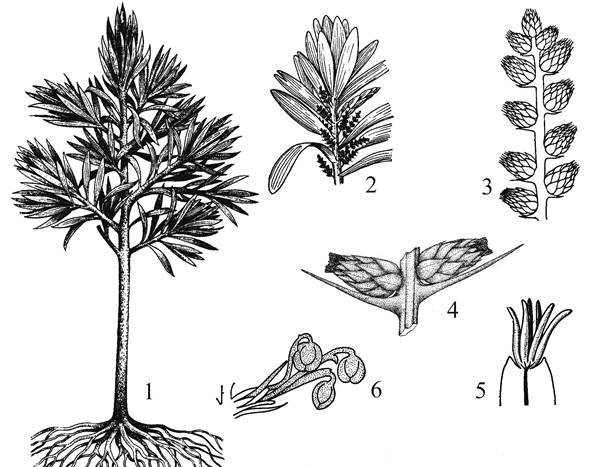
Рис. 15. Кордаитовые: 1 – общий облик кордаита (реконструкция); 2 – ветвь с собраниями микростробилов; 3 – собрание микростробилов; 4 – фрагмент оси с двумя пазушными микростробилами; 5 – верхушка микроспорофилла с микроспорангиями; 6 – фрагмент мегастробила с тремя семяпочками на укороченном пазушном побеге.
У кордаиантуса изучено строение семяпочки. Интегумент состоял из 3 слоев, был свободный, не сросшийся с нуцеллусом до основания, снабжен проводящим пучком, который разделяется на два; В нуцеллусе имеется лопастная пыльцевая камера и выступающий в нее клювик. Клеточной ткани в семяпочке гаметофита обычно не обнаруживается. Это может быть связано с тем, что гаметофит имел свободноядерную стадию в своем развитии и полностью разрушался при захоронении. Семена с крыловидными выростами, плоские.
Описаны также своеобразные зонтиковидные мегастробилы (Gaussia), кистевидные с укороченной или удлиненной осью стробила и разной длиной семяножки (Krylovia).
Изучение строения сложных мегастробилов разного типа имело большое значение для выяснения вопросов, связанных с происхождением сложной шишки – констробила (типа сосновых) и морфологической природы составляющих ее структур – семенной и кроющей чешуй.
Окаменевшие остатки кордаитовых дали мощные залежи каменного угля (например, в Кузбассе).
Подкласс Хвойные, Пиниды (Pinidae)
Хвойные являются наиболее многочисленной группой современных голосеменных растений. По жизненным формам среди них можно выделить такие: деревья с моноподиальным типом ветвления с прямостоячим стеблем; кустарники с прямостоячими либо стелющимися стеблями, называемые стланиками. Род Aethophyllum из вольтциевых (вымершая группа) включал, предположительно, травянистые растения. Древесным паразитным растением является представитель семейства подокарповых – паразитаксус опаленный, который представляет собой невысокий ветвящийся кустарник, корни которого проникают в корни и стволы другого представителя подокарповых – фалькатифолиума тиссолистного (Новая Каледония).
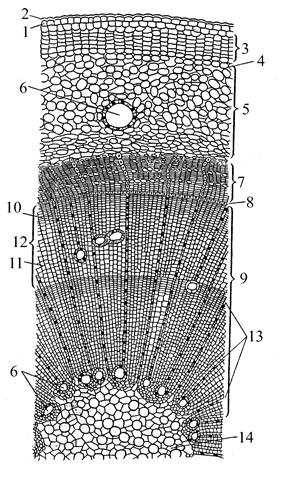
По особенностям анатомического строения хвойные относятся к пикноксильным древесным растениям, т. е. имеющим плотную, мощную древесину, относительно тонкую кору и слабо выраженную сердцевину.
Ксилема у хвойных на 90-95% образована трахеидами. В протоксилеме трахеиды с кольчатыми и спиральными утолщениями, в метаксилеме и вторичной ксилеме чаще всего трахеиды с окаймленными округлыми, реже шестигранными порами с торусом, расположенными на радиальных стенках. У араукариевых торус не всегда выражен. У тисса и головчатотисса имеются еще спиральные утолщения в виде 1-2 лент, которые являются третичными образованиями проводящих элементов ксилемы. У хвойных умеренной зоны в древесине обычно хорошо выражены годичные кольца.
 Древесинная паренхима у хвойных слабо представлена либо вовсе отсутствует. Она располагается обычно вокруг смоляных ходов, реже между трахеидами.
Древесинная паренхима у хвойных слабо представлена либо вовсе отсутствует. Она располагается обычно вокруг смоляных ходов, реже между трахеидами.
Сердцевинные лучи обычно 1-2-рядные, состоят из паренхимных клеток, иногда с краевыми рядами поперечно расположенных лучевых трахеид (рис. 16).
У многих хвойных имеются смоляные ходы в древесине и в коре постоянно, а у ряда представителей они образуются только при повреждениях (травматические смоляные каналы). У подокарповых смоляные каналы есть только в листьях. У араукариевых смола содержится в трахеидах, окаймляющих лучи. Флоэма хвойных состоит из ситовидных клеток с клетками Страссбургера. С началом вторичного роста в толщину эпидермис замещается перидермой, которая закладывается во внешнем слое паренхимы первичной коры.
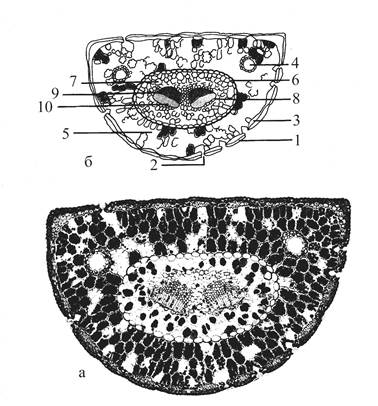
| Рис. 16. Поперечный срез стебля сосны. 1 – эпидермис с кутикулой (2); 3 - пробка; 4 – пробковый камбий; 5 – первичная кора; 6 – смоляные ходы; 7 – флоэма; 8 – камбий; 9 – ксилема; 10 – осенняя древесина; 11 - весенняя древесина; 12 – годичное кольцо; 13 – сердцевинные лучи; 14 – сердцевина. |
У хвойных листья располагаются по-разному. У многих видов они находятся на обычных удлиненных побегах, размещаясь по спирали, супротивно, реже мутовчато. Для ряда хвойных характерно наличие укороченных побегов (брахибластов), имеющих ограниченный прирост. Они обычно несут на верхушке пучок сближенных зеленых листьев, число которых бывает строго определенным (у сосен 2-3-5), или неопределенно большим (у лиственницы, кедра 20-50). Часто у основания укороченных побегов имеются рано опадающие чешуйчатые листья. У отдельных представителей зеленые листья располагаются как на укороченных, так и на удлиненных побегах.
Ежегодный листопад происходит у немногих хвойных (лиственница, лжелиственница). Большинство пинид – вечнозеленые растения. У некоторых наблюдается веткопад (таксодиум мексиканский, метасеквойя), опадают удлиненные побеги с листьями. У некоторых сосен пучок листьев опадает вместе с укороченным побегом.
 Листья хвойных по форме отличаются некоторым разнообразием: игловидные, чешуевидные, линейные, ланцетные. В поперечном сечении они бывают плоскими, трехгранными, четырехгранными. По расположению устьиц на поверхности листа различают: амфистомные (устьица на верхней и нижней поверхности), гипо- или эпистомные, причем в онтогенезе первые листья чаще амфистомные. У филлокладуса роль листьев выполняют филлокладии.
Листья хвойных по форме отличаются некоторым разнообразием: игловидные, чешуевидные, линейные, ланцетные. В поперечном сечении они бывают плоскими, трехгранными, четырехгранными. По расположению устьиц на поверхности листа различают: амфистомные (устьица на верхней и нижней поверхности), гипо- или эпистомные, причем в онтогенезе первые листья чаще амфистомные. У филлокладуса роль листьев выполняют филлокладии.
| Рис. 17. Внутренне строение хвои сосны: а – поперечный срез; б – схема строения. 1 – эпидермис; 2 – устьице; 3 – гиподерма; 4 – смоляной ход; 5 – мезофилл; 6 – эндодерма; 7 – трансфуззионная ткань; 8 - проводящий пучок; 9 – ксилема; 10 – флоэма. |
Листовые следы у большинства хвойных двойные, у некоторых – простые. Реже в листе имеется несколько жилок, которые затем могут дихотомически ветвиться у основания (агатис, подокарп).
По анатомическому строению листья хвойных имеют некоторые различия у ксерофитов и мезофитов. Так, ксероморфные игловидные листья с толстым слоем кутикулы поверх эпидермиса, у них погруженные устьица, слой толстостенной гиподермы и губчатая ткань мезофилла. Одни или два проводящих пучка окружены особой трансфузионной тканью, в которой имеются живые паренхимные клетки и короткие трахеиды. Эта ткань обеспечивает связь мезофилла с проводящими тканями. К ней примыкает слой эндодермы. В мезофилле листа проходят смоляные каналы (рис. 17).
Листья некоторых хвойных с широкой плоской листовой пластинкой (араукариевые, подокарповые) имеют разветвленную сеть проводящих пучков, мезофилл состоит из палисадной и губчатой паренхимы. Камбий в листовых пучках прекращает свою деятельность в первый год жизни листа.
Корневая система большинства хвойных с хорошо выраженным главным корнем и разветвленными боковыми. У многих представителей образуется микориза, а у подокарповых корни вступают в симбиоз с азотфиксирующими бактериями, образуя клубеньки, наподобие таковых у бобовых.
Корневые волоски у хвойных располагаются на корнях на небольшом участке вблизи верхушки корня. Протоксилема корня экзархная, имеется многослойный перицикл и 1-слойная эндодерма.
У большинства хвойных репродуктивные структуры представляют собой мужские (микростробилы) и женские шишки (сложные мегастробилы, констробилы). По общему плану строения и по отдельным элементам имеются различия, которые характеризуют те или иные таксоны хвойных и рассматриваются при характеристике семейств.
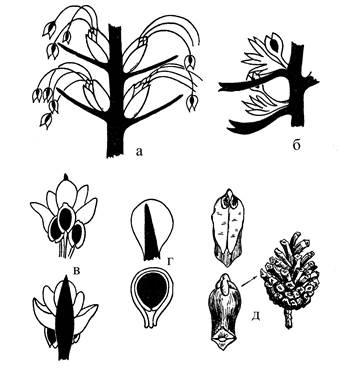
Сосна обыкновенная – растение однодомное. Мужские шишки располагаются группами на молодых побегах и занимают боковое положение на некотором расстоянии от верхушки, которая продолжает рост побега. Отдельная мужская шишка длиной около 1 см, она представляет собой простой микростробил. На оси шишки расположены пленчатые микроспорофиллы, несущие по два микроспорангия в виде продолговатых пыльцевых мешков на нижней поверхности микроспорофилла. В них из микроспор начинают развиваться мужской гаметофит – пыльцевое зерно. Сформировавшееся пыльцевое зерно с 2-мя воздушными мешками переносится на семяпочку ветром (рис. 2). Заложение мужской шишки происходит осенью, а созревание пыльцы – весной.
Женская шишка (семенная шишка) образуется на верхушке удлиненного побега, по сравнению с мужской шишкой она имеет более сложное строение. В первый год жизни она бывает красноватого цвета и достигает в длину 6-8 мм. На оси шишки очень компактно расположены плотные дорзовентральные семенные чешуи, на верхней стороне которых ближе к основанию находится по две семяпочки.
Каждая семенная чешуя расположена в пазухе тонкой пленчатой кроющей чешуи, которая короче семенной, поэтому видна только с нижней стороны (рис. 2). И в кроющую, и в семенную чешуи входят проводящие пучки, в которых взаимное расположение ксилемы и флоэмы показывает различную природу этих образований.
| Рис. 18. Происхождение женской шишки сосны. Фрагмент собрания мегастробилов кордаита (а) и лебахии (б); пазушный комплекс псевдовольтции (в); семенная и кроющая чешуи ульмании (г); женская шишка сосны (д). |
 Относительно морфологической природы и происхождения семенной чешуи высказывалось много различных предположений. Однако в настоящее время можно считать, пожалуй, общепризнанным, что семенная чешуя не является мегаспорофиллом, т. е. не имеет листовой природы, а представляет собой видоизмененный мегастробил и в основе своей имеет побеговую природу, о чем свидетельствует анатомическое строение. Таким образом, вся женская шишка сосны представляет собой скученное собрание мегастробилов, или констробил (рис. 18).
Относительно морфологической природы и происхождения семенной чешуи высказывалось много различных предположений. Однако в настоящее время можно считать, пожалуй, общепризнанным, что семенная чешуя не является мегаспорофиллом, т. е. не имеет листовой природы, а представляет собой видоизмененный мегастробил и в основе своей имеет побеговую природу, о чем свидетельствует анатомическое строение. Таким образом, вся женская шишка сосны представляет собой скученное собрание мегастробилов, или констробил (рис. 18).
Семяпочка сосны обращена пыльцевходом к основанию семенной чешуи. Интегумент на всем протяжении, кроме верхушки, срастается с нуцеллусом. Развитие семяпочки у сосны протекает очень медленно. Рано весной в семяпочке первого года жизни еще неразличима спорогенная ткань. В конце весны на микропилярном полюсе нуцеллуса образуется обычно одна функционирующая материнская клетка мегаспор (мегаспороцит). В результате мейотического деления возникает 4 мегаспоры, расположенные линейно в направлении длинной оси семяпочки. В дальнейшем только самая нижняя мегаспора дает начало женскому гаметофиту, остальные дегенерируют.
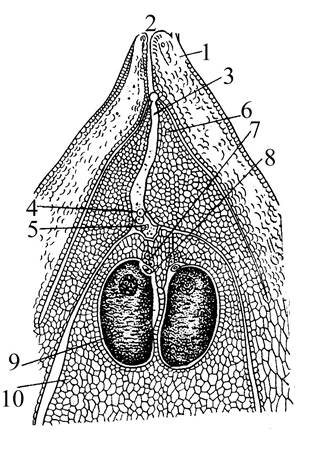 Для начальной стадии формирования женского гаметофита характерно свободноядерное деление (до 2000 ядер), и сплошное расположение слоя цитоплазмы вокруг образующейся крупной центральной вакуоли. Затем происходит обособление цитоплазмы вокруг ядер, формирование первичных стенок и увеличение объема клеток, интенсивное отложение в них запасных питательных веществ. Формирование клеток происходит по направлению от периферии к центру женского гаметофита.
Для начальной стадии формирования женского гаметофита характерно свободноядерное деление (до 2000 ядер), и сплошное расположение слоя цитоплазмы вокруг образующейся крупной центральной вакуоли. Затем происходит обособление цитоплазмы вокруг ядер, формирование первичных стенок и увеличение объема клеток, интенсивное отложение в них запасных питательных веществ. Формирование клеток происходит по направлению от периферии к центру женского гаметофита.
|
|