Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Для новых пользователей первый месяц бесплатно. Чат-бот для мастеров и специалистов, который упрощает ведение записей: — Сам записывает клиентов и напоминает им о визите;
— Персонализирует скидки, чаевые, кэшбэк и предоплаты;
— Увеличивает доходимость и помогает больше зарабатывать; Начать пользоваться сервисом

Часть 1. Некоторые сведения о физиологии нервных клеток.
|
|
ТЕМА 13. гАЗООБРАЗНЫЕ НЕЙРОМОДУЛЯТОРЫ И ОБЪЕМНАЯ ПЕРЕДАЧА СИГНАЛА
Часть 1. Некоторые сведения о физиологии нервных клеток.
- Концепции объемной и проводниковой передачи сигнала.
- Клеточная организация мозга.
- Типы межклеточных сигналов
- Кислотно-основное равновесие (pH) и температура как факторы объемной передачи сигнала.
Часть 2. Газообразные нейромодуляторы.
- Монооксид азота (NO).
- Монооксид углерода (СO) и сульфид водорода (Н2S).
Часть 1. Некоторые сведения о физиологии нервных клеток.
- Концепции объемной и проводниковой передачи сигнала.
Разделение процессов межклеточной коммуникации на основании особенностей распространения и восприятия передаваемого сигнала было впервые проведено Л. Агнати в 1986 году.
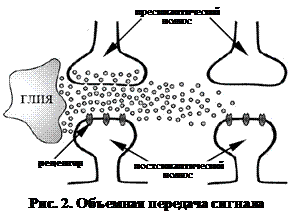
Согласно высказанной ими концепции, выделяют следующие способы передачи сигнала (рис. 1, 2):
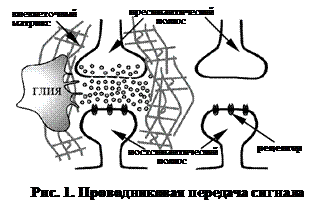 Проводниковая передача сигнала – способ межклеточной коммуникации, опосредованный детерминированной цепочкой клеток. При этом наблюдается передача сигнала от одной клетки к другой с коэффициентом 1: 1, т. е. на один источник сигнала (пресинаптическую клетку) приходится один приемник этого сигнала (постсинаптическая клетка). Представлена классической синаптической передачей, например в случаях простых рефлекторных дуг соматических рефлексов, хотя наличие в них вставочных нейронов несколько и не соответствует декларированному коэффициенту 1: 1.
Проводниковая передача сигнала – способ межклеточной коммуникации, опосредованный детерминированной цепочкой клеток. При этом наблюдается передача сигнала от одной клетки к другой с коэффициентом 1: 1, т. е. на один источник сигнала (пресинаптическую клетку) приходится один приемник этого сигнала (постсинаптическая клетка). Представлена классической синаптической передачей, например в случаях простых рефлекторных дуг соматических рефлексов, хотя наличие в них вставочных нейронов несколько и не соответствует декларированному коэффициенту 1: 1.
Объемная передача сигнала – способ межклеточной коммуникации, основанный на распространении сигнала в пределах межклеточного пространства во всех направлениях. При этом расстояние, на которое такой сигнал распространяется, значительно превышает размеры синаптической щели. Во внеклеточном пространстве сигнал распространяется по пути, обеспечивающему наибольшую скорость диффузии сигнальной молекулы. Передача сигнала происходит с коэффициентом 1: n, где n> > 1. Таким образом, количество приемников сигнала значительно превышает количество источников исходного сигнала.
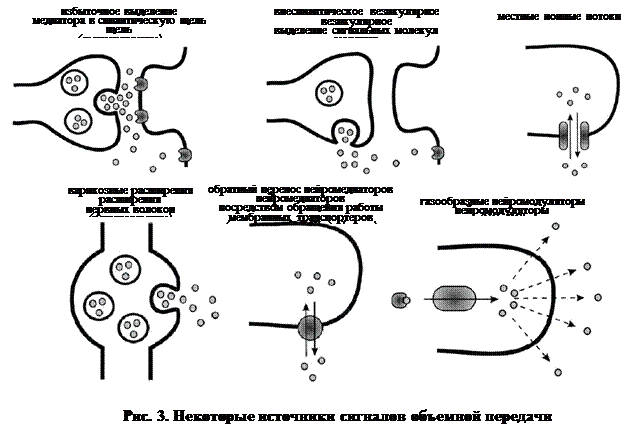
Многие нейромедиаторы и нейромодуляторы способны выступать в качестве сигнальных молекул объемной передачи сигнала (рис. 3), однако со всей очевидностью эта роль наиболее характерна для газообразных нейромодуляторов. Разумеется, что клетки-мишени объемной передачи сигнала обладают специфическими молекулами, способными воспринимать и распознавать исходный сигнал (рецепторы).
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение
В современной нейробиологии уже не вызывает сомнений, что в мозге, помимо синаптической передачи, существует и другая, гормон-подобная передача сигнала. Основу такого паракринного влияния составляет генерализованное действие того или иного вещества.
При этом в ответную реакцию вовлекается целая совокупность клеток, расположенных на некотором расстоянии от клетки, источника сигнала. Наряду с газообразными нейромодуляторами, многие нейропептиды, а также и белковые факторы роста относятся к паракринно действующим началам.
Помимо межклеточного пространства существует и другой путь, по которому сигналы могут распространяться от клетки к клетке по всем направлениям. Это внутриклеточное пространство расположенных рядом нейронов, соединенных посредством щелевого контакта. В отличие от первого (внеклеточного) пути, он представляет собой хорошо регулируемый путь, позволяющий координировать работу функционально однородных нейронов. Так, известно о том, что синтезируемый амакриновыми клетками сетчатки глицин может поступать в связанные с ними биполярные клетки через щелевой контакт.
- Клеточная организация мозга.
Несмотря на чрезвычайную сложность клеточной организации мозга, все разнообразие нейронных сетей можно свести к двум базовым схемам:
1. Иерархические системы – к ним относятся многочисленные проводящие пути спинного и головного мозга, контролирующие двигательную активность и чувствительность и состоящие из различных клеток последовательно соединенных при помощи толстых, миелинизированных волокон. Важной особенностью этой схемы является то, что повреждения, нанесенные на любом ее уровне, вызывают нарушение работы всей системы.
Иерархические системы состоят из клеток двух типов:
ü проекционные (релейные) нейроны: будучи связанными посредством химического синапса, передают сигналы на большие расстояния в пределах организма.
Для них характерны крупные тела с многочисленными отростками вокруг (дендриты) и одним длинным отростком (аксон) для контакта с другими составляющими системы. Как правило, синапсы, образованные этими клетками, относятся к возбуждающим синапсам, часто использующим глутамат в качестве нейромедиатора;
ü вставочные (местные) нейроны: они участвуют в образовании многочисленных синапсов с проекционными нейронами или друг с другом.
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
Тела этих клеток значительно мельче, аксоны мало выражены. Разнообразие синапсов (аксо-соматические, аксо-дендритные, аксо-сомальные и т. п.) позволяет реализовать различные виды межнейронных взаимодействий, прежде всего торможение – прямое, возвратное, латеральное и др.
Таким образом, большинство синапсов, образованных вставочными нейронами, – тормозные, предпочтительно использующие ГАМК и/или глицин в качестве нейромедиатора.
Несмотря на разнообразие синаптических связей, в подсистеме вставочных нейронов используется ограниченное количество нейромедиаторов. В результате, фармакологически воздействуя на них, можно существенно изменить общую возбудимость нервной системы, что и наблюдается, например, при действии стрихнина.
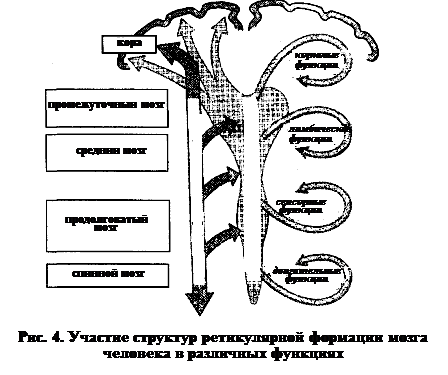
2. Диффузные (неспецифические) системы – к ним относятся нейронные сети ретикулярной формации мозга. При этом тела клеток входят в состав относительно немногочисленных (в мозге человека насчитывается около 40 ядер) компактных скоплений. Отростки нейронов ретикулярной формации тонкие, немиелинизированные, часто ветвятся и меняют направление, следуя как в восходящем, так ив нисходящем направлении. На нейронах ретикулярной формации конвергируют различные афферентные сигналы, в то же время ветви одного нейрона могут иннервировать несколько функционально различных частей ЦНС.
В качестве нейромедиаторов диффузные системы используют различные моноамины – норадреналин, серотонин и, главное, дофамин.
На протяжении аксонов моноаминергических нейронов существуют многочисленные расширения (аксоны en passant), содержащие большое количество синаптических пузырьков, обеспечивающих диффузное выделение нейромедиатора. Важным следствием подобной организации является то, что клеточные мишени действия медиатора определяются местоположением их рецепторов, а не источника их синтеза.
В связи с этим диффузные системы не могут и не являются передатчиками специфической топографической информации. Они опосредуют генерализованные восходящие тонические влияния (возбуждающие и тормозные), направленные на регуляцию активности коры больших полушарий, вовлечены в регуляцию процессов сна и бодрствования, эмоциональных состояний.
- Типы межклеточных сигналов
В развитие концепции объемной передачи сигнала было предложено разделить всю совокупность сигналов, воспринимаемых клетками нервной системы, на два класса:
1. Специфические сигналы – для их распознавания в клетке должны существовать специальные структуры, взаимодействующие с ними за счет высокоаффинного связывания, т. е. рецепторы. В эту группу входят нейромедиаторы и нейромодуляторы, высвобождаемые ограниченным количеством групп нервных клеток.
2. Общедоступные сигналы – генерируются и распознаются (декодируются) всеми нервными клетками, например, электрические сигналы.
Заметим, что такие общебиологические факторы, как температура, вне- и внутриклеточная концентрация Н+ (pH), способны оказывать модулирующее влияние на процессы генерации сигнала и на процессы его восприятия и распознавания.
Более того, указанные физические и физико-химические начала с легкостью могут быть отнесены к группе общедоступных сигналов в самом широком понимании, поскольку практически любая клетка способна реагировать на изменение температуры или pH.
- Кислотно-основное равновесие (pH) и температура как факторы объемной передачи сигнала.
В ЦНС тканевый ацидоз является одним из следствий ишемии мозга или развития различных воспалительных процессов, а то, что даже малые сдвиги уровня pH способны вызвать выраженные изменения возбудимости нервных клеток, не вызывает сомнений.
В ходе тканевого ацидоза значение внеклеточного pH может снижаться более чем на две единицы по сравнению с исходным уровнем.
Помимо этого, сообщалось о модулирующем влиянии уровня pHна электрофизиологические характеристики Са2+-токов, а следовательно, и функционирование кальциевых каналов, ГАМК-управляемых хлорных каналов и др. Совершенно очевидно, что эти данные свидетельствуют о вовлеченности синаптических механизмов (пре- и постсинаптических составляющих) в ответную реакцию организма на сдвиги внеклеточного pH. В частности, падение pH подавляет дофамин- и пептидергическую передачу в нервной системе беспозвоночных.
Изменение величины pH, а особенно закисление внеклеточной среды, приводит к активации открытых в начале 80-х гг. XX в. кислоточувствительных ионных каналов, широко представленных в сенсорных нейронах спинно-мозговых ганглиев, нейронах ЦНС и клетках глии (олигодендроциты). Падение pH приводит к открытию этих неселективных катионных каналов (Na+ > Ca2+ > K+), изменяет возбудимость нейронной мембраны, что и опосредует реакцию соответствующей клетки.
Физиологическая роль кислородчувствительных ионных каналов остается до конца не выясненной, хотя не вызывает сомнений их вовлеченность в процессы, связанные с болевой чувствительностью. Возможно, сдвиги pH внутренней среды организма играют регуляторную роль, неодинаково модулируя работу функционально различных нейронных сетей и, как следствие, контролируемых ими форм поведенческой активности.
По-видимому, протон – это самый «простой» лиганд, вовлеченный в процессы межклеточной коммуникации, выступающий, по меньшей мере, в качестве нейромодулятора.
Абиотические внешние факторы, например температура, также способны радикально изменить протекание процессов межклеточной коммуникации, прежде всего синаптической передачи. Очевидно, что любой из этапов химической передачи сигнала подвержен модулирующему влиянию температуры.
В то же время различные нейромедиаторные системы демонстрируют неодинаковую температурную зависимость эффективности синаптической передачи.
Проницаемость щелевых контактов также подвержена действию температурного фактора.
Таким образом, генерализованные влияния факторов объемной передачи сигнала реализуются посредством одномоментного вовлечения в ответную реакцию множества компонентов нейронных сетей.
|
|