Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника

Раздел Amniota. Высшие позвоночные
|
|
К высшим позвоночным относятся парарептилии, рептилии, птицы и млекопитающие. От низших позвоночных эти группы отличаются наличием у эмбриона зародышевых оболочек (амнион и аллонтоис), при помощи которых осуществляется его дыхание и питание. Зародышевые оболочки позволяют размножаться вне водной среды без личиночной стадии и метаморфизма. В некоторых классификациях всю группу называют амниотами (Amniota).
Парарептилии и рептилии ранее рассматривались в составе одного класса рептилий. Сейчас парарептилии обычно выделяют в отдельный класс, хотя это подразделение и является достаточно искусственным. Классификации парарептилий дискуссионны.
Класс Parareptilia. Парарептилии
В класс объединяют представителей четвероногих, сочетающих признаки двух классов – земноводных и рептилий. Они размножаются яйцами, имеют зародышевые оболочки и пятипалые конечности, легочное дыхание (жаберное редко), чем сближаются с рептилиями. В то же время они имеют некоторые признаки земноводных: у них отсутствуют височные окна, у ряда форм развита сплошная крыша черепа из скульптированных покровных костей, имеется теменное отверстие и пр.
Образ жизни парарептилий разнообразный. Поскольку большинство из них дышит легкими, они обитают на суше или в пограничных водно-прибрежных зонах. Есть также вторично-водные формы. Большинство парарептилий – растительноядные, хотя есть и всеядные и хищники. Поздний карбон – современность. В класс входят несколько групп животных, из них наиболее известны котилозавры и черепахи.
Подкласс Котилозавры. Cotylosauria.. C3-T. Это первые настоящие наземные позвоночные, которые дали начало остальным подклассам рептилий. Они появились в позднем карбоне, а вымерли в триасе. Котилозавры (рис. 22) произошли от амфибий – батрахозавров, с которыми схожи строением черепа.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз, а первые результаты появляются уже в течение первых 7 дней. Зарегистрироваться и Начать продвижение

Рис. 22. Котилозавр лабидозавр (Labidosaurus, P1) [6]
У котилозавров сохранился панцирь на голове. Череп не имел височные впадины. Такой тип черепа (с покровными окостенениями и без височных впадин) называется анапсидным. Череп скульптирован. Морда короткая, реже умеренно удлиненная. Конечности были короткими и массивными.
Группа включает как мелких наземных ящерицеобразных животных, так и крупных растительноядных, величиной с бегемота.
Известный представитель котилозавров – парейазавр, или щекастый ящер (рис. 23, 24) – неуклюжее грузное животное до 3 м длиной. Череп широкий большой скульптированный с массивными выступами («щеками») по бокам, морда округлая, зубы долотовидные. Нередко был спинной панцирь из мозаично расположенных костных пластинок. Растительноядный.

Рис. 23. Котилозавр – парейазавр (Pareiasaurus, P) (реконструкция) [23]
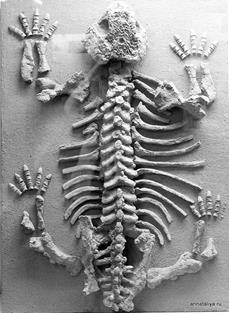
Рис. 24. Скелет молодого парейазавра [47]
Котилозавры вели различный образ жизни, по способу питания среди них были хищники, растительноядные, моллюскоядные, насекомоядные. Остатки котилозавров многочисленны в пестроцветных фациях дельт и аллювиальных равнин. Животные используются в стратиграфии перми, нижнего и среднего триаса. Характерная форма: парейазавр - Pareiasaurus (P2-3).
Подкласс Черепахи. Chelonia. Считается, что черепахи произошли от котилозавров в перми, однако остатки их первых представителей нашли только в верхнем триасе. Эти черепахи уже обладали почти всеми характерными признаками группы. Но во рту у них присутствовали зубы. У более молодых форм черепах имелся роговой клюв.
В подкласс объединяют животных с коротким и широким туловищем, заключенным в костный панцирь. Его и находят в ископаемом состоянии (рис. 25). Размеры черепах различны. Одна из самых крупных черепах, архелон, была найдена в меловом периоде. Размер ее скелета достигал 4 м. Черепахи, почти не изменившись, дожили до наших дней.

Рис. 25. Скелет черепахи (Triassochelys, Т1) [6]
В геологии черепахи используются в основном как показатели климата прошлых эпох. Так, нахождение речных и озерных черепах указывает на влажный и теплый климат. Наличие роющих черепах с уплощенным панцирем свидетельствует об открытых степных или пустынных ландшафтах. Некоторые черепахи имеют значение в стратиграфии палеогена и неогена.
2.2 Класс Reptilia. Пресмыкающиеся (Рептилии)
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно. Зарегистрироваться в сервисе
Ранние рептилии имели общие черты строения с земноводными и парарептилиями, более позние – с птицами и млекопитающими (появление теплокровности, вторичного неба, четырехкамерного сердца, перьев, шерсти и пр.).
От парарептилий рептилии отличаются, прежде всего, строение черепа: сзади глазниц он не является сплошным, а несет одно или два отверстия – височные окна (ямы).
От земноводных рептилии отличаются следующими основными признаками:
1. Размножением яйцами, имеющими кроме зародышевой оболочки также твердую скорлупу, что в совокупности явилось исключительно важным приспособлением позвоночных к наземной жизни. Из яйца появляется молодое животное, которое отличается от взрослого только размерами.
2. Развитием легочного дыхания, что привело к необходимости защитить голую кожу животного покрытием (роговые чешуи, костные пластинки). Это явилось вторым важным шагом на пути приспособления к сухопутной жизни.
3. Разнообразием условий существования (земля, вода, воздух).
4. Более совершенными и сложными кровообращением и выделительной системой. В процессе дальнейшего развития центральной нервной системы появилась кора мозга в больших полушариях.
5. Значительными изменениями в строении скелета. В височной области появляются височные впадины, служащие вместилищем челюстной мускулатуры (более сильной, чем у амфибий). Позвоночник приобретает четко выраженную дифференцировку: в частности, обособляется шейный отдел (у земноводных отсутствовал). Это дало животному свободу движений. В теле позвонков постепенно исчезает отверстие для хорды. Парные конечности стали разнообразнее и совершеннее: развились суставные поверхности, конечности удлинились. Это позволило поднять тело высоко, не волоча его по земле, что было необходимо для быстрого передвижения.
Вместе с тем у рептилий сохранились и ряд примитивных признаков: плохо развиты органы чувств (кроме зрения), слуховой аппарат (за небольшим исключением) состоит из одной косточки, т.е. как у амфибий, зубы у многих не дифференцированы и т.д. Как и земноводные, большинство пресмыкающихся являются холоднокровными животными с непостоянной температурой тела.
Рептилии появляются в середине карбона, в мезозое широко распространены, к концу мезозоя большая часть их вымирает. Современные представители класса – змеи, ящерицы, крокодилы, гаттерии и др. В ископаемом состоянии встречаются как полные скелеты рептилий, так и их части. Довольно часты следы лап рептилий. Их можно найти в местах захоронения скелетов животных, или отдельно. Следы очень разнообразны по облику, размерам и сохранности. Обычны копролиты. Наиболее крупные скопления остатков рептилий наблюдаются в области дельт, т.е. там, где были самые благоприятные условия для захоронения: концентрация трупов течением и их быстрое занесение осадком. В озерах же, наоборот, встречаются обычно отдельные скелеты и их части.
С биологической точки зрения значение пресмыкающихся и парарептилий в истории развития органического мира нашей планеты велико: именно они являются непосредственным промежуточным звеном между амфибиями, птицами и млекопитающими. Родственные связи прослеживаются через котилозавров с амфибиями, через архозавров – с птицами, через зверообразных – с млекопитающими.
Геологическое значение рептилий также велико. Сухопутные рептилии (котилозавры, зверообразные, динозавры) широко используются в стратиграфии верхнепалеозойских и мезозойских, в основном континентальных отложений. Остатки морских пресмыкающихся (ихтиозавров, плезиозавров, мозазавров) имеют меньшую ценность из-за их редкости, однако при их обнаружении окаменелости применяют в основном для расчленения верхнеюрских и меловых отложений.
Класс Пресмыкающихся объединяет пять подклассов: Зверообразные –Synapsida (С3-К1), Чешуйчатые – Lepidosauria (Р2-Q), Эвриапсиды – Euryapsida (Т-K), Ихтиозавры или Рыбоящеры – Ichthyopterygia (T-K2), Архозавры – Archosauria (Р2-Q).
Подкласс Зверообразные. Synapsida. Эти рептилии имели синапсидный (одно височное окно) тип черепа. В ходе эволюции приобрели черты млекопитающих: увеличилась зубная кость нижней челюсти, появилась дифференциация зубов (резцы, клыки и коренные зубы), появился шерстистый покров и теплокровность и т.д. Череп зверообразных был высоким и узким. Конечности не отличались по длине или задние были длиннее передних. Многие имели почти прямую постановку конечностей. Большинство синапсид - хищники. Жили со среднего карбона – по среднюю юру.
В конце карбона они возникли от котилозавров, в свою очередь, их представители в триасе дали начало млекопитающим. Примером зверообразных является иностранцевия - Inostrancevia (рис. 30, 31), жившая в перми. Рептилия имела дифференцированные зубы: резцы, заклыковые зубы, мощные клыки, а на пальцах – большие когти. Животное достигало 3 м в длину при длине черепа 50 см.
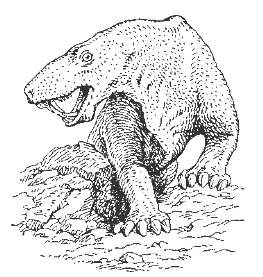 Рис. 26. Реконструкция рептилии
Рис. 26. Реконструкция рептилии
подкласса зверообразных – иностранцевии (Inostrancevia, P2) [1]
Интересна также двиния (Dvinia) из поздней перми – мелкое всеядное животное с шерстистым покровом, похожее на крысу (рис. 27). Двинии относятся к последнему и самому продвинутому подотряду зверообразных – цинодонтам – прямым предкам млекопитающих. Они уже были полностью покрыты шерстью и мало отличались от первых триасовых млекопитающих. Есть мнение, что современные утконос и ехидна – на самом деле цинодонты, сохранившиеся до наших дней.

Рис. 27. Dvinia – представитель зверообразных –
потомков млекопитающих [45]
Характерные представители зверообразных: иностранцевия – Inostrancevia (P2-3), двиния – Dvinia (P3).
Подкласс Lepidosauria. Чешуйчатые. Это примитивные ящерицеобразные пресмыкающиеся (чешуйчатые ящеры), известные с перми. В палеозое они имели диапсидный тип черепа (две височные дуги и два окна). В течение мезозоя и кайнозоя дуги и окна частично или полностью исчезли.
У чашуйчатых были парные конечности в основном умеренной длины, но у змей и некоторых ящериц они исчезают, хотя рудименты поясов конечностей могут сохраняться. У некоторых водных форм конечности становятся ластовидными. У лепидозавров сохраняется обычный для ранних четвероногих примитивный тип передвижения с волнообразным изгибанием туловища.
Лепидозавры жили в разных услових (морях, болотах, озерах, на суше и т.д.), поэтому их остатки встречают в разных породах. Стратиграфическое значение лепидозавров невелико. Но те из них, которые имеют узкий диапазон распространения, например, мозозавры, с успехом используются для расчленения мезозойских отложения.
Вымершие чешуйчатые представлены наземными ящерицеобразными эозухиями и хищными морскими ящерицами – мозозаврами. Они имеют диапсидный тип черепа и являются переходной группой между парарептилиями – котилозаврами и прогрессивными рептилиями – архозаврами. Современные представители подкласса – змеи, ящерицы, хамелеоны, гаттерии. Змеи обособились от ящериц в конце мела.
Эозухии (рис. 28) внешне были похожи на ящериц или крокодилов. Первые имели небольшие размеры – до 50 см, крокодилообразные могли достигать 7, 5 м. От настоящих ящериц и крокодилов отличались строением черепа. Они появились в перми, в триасе дали начало другим чешуйчатым: клювоголовым и ящерицам. Поздний карбон – ранний триас.


Рис. 28. Эозухии: а – скелет [6], б – реконструкция [39]
Мозозавры (рис. 29) – морские ящерицы, жившие в позднем мелу, имели удлиненное тело до 15 м в длину, крупную голову и две пары пятипалых конечностей – ласт, хвост превращен в плавник, который являлся главным органом движения.

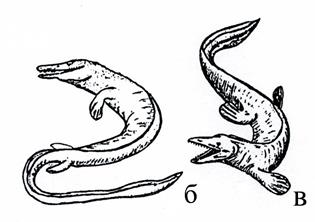
| Рис. 29. Мозозавры – морские ящерицы: а – скелет [6], б-в – реконструкция животных |
Подкласс Эвриапсиды (Euryapsida) или Synaptosauria. Синаптозавры.. Эти пресмыкающиеся вели водный (морской) образ жизни и все являются плотоядними. Они имели обычно крупное, широкое (боченковидное) туловище, короткий хвост и часто длинную шею. Длина тела достигала 16 м. Тип черепа эвриапсидный, характеризующийся одной верхней височной ямой. Конечности – ластовидные.
Синаптозавры представлены двумя отрядами. Представители одного из них (плезиозавры, плиозавры и др.) имели конические острые зубы (T-K), другого – уплощенные зубы (плакодонты) (T2-J1).
Плезиозавры (рис. 30) – животные с маленькой головой, длинной, гибкой шеей и коротким хвостом. Длина до 16 м. Это активные хищники. Обитали вблизи берега, питались рыбой. Классические местонахождения плезиозавров находятся в Англии. Триас-мел.
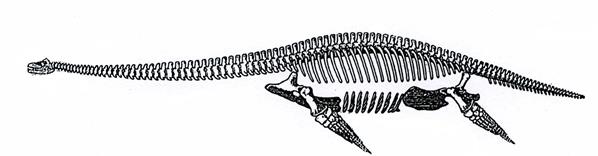
Рис. 30. Плезиозавр – муренозавр (Muraenosaurus, J3) [6]

Рис. 31. Плезиозавры (реконструкция) [14]
Плиозавры (рис. 32) – хищники огромных размеров с короткой шеей и крупной головой (череп до 4 м в длину), с сильными коническими зубами и хвостом разной длины. Длина тела до 15 м. Обитатели открытых морей в триасе и мелу. Принадлежали к числу опаснейших морских хищников своего времени.


Рис. 32. Плиозавры: а – череп плиозавра сбоку [6], б – реконструкция [40].
Плакодонты (рис. 33) – морские пресмыкающиеся, часть которых похожа на черепах. Они имели уплощенное (иногда удлиненное) тело, защищенное костными щитками, иногда образующими полный панцирь. Шея короткая и толстая. Конечности ластовидные. Те плакодонты, которые имели острые зубы, являлись малоподвижными хищниками морского мелководья, они питались моллюсками и брахиоподами. Те, у которых вместо зубов имеется клюв, питались планктоном и водорослями. Длина плакодонтов достигала 2, 5 м. Могли некоторое время проводить на суше. Средний триас – ранняя юра.
Есть мнение, что плезиозавры, плиозавры и плакодонты возникли от котилозавров. Согласно другой точке зрения, синаптозавры берут свое начало от примитивных диапсид, вероятно, близким эозухиям (см. ниже).

| 
|
Рис. 33. Плакодонты: а – скелет (Placochelys, T3) [6], б – реконструкция [20]
Остатки синаптозавров встречаются редко, поэтому стратиграфическое значение их невелико. Тем не менее, плезиозавры и плиозавры входят в состав типовых комплексов позвоночных верхнего триаса-нижнего мела и используются для расчленения и определения возраста отложений этого уровня. Плакодонты, имеющие узкое распространение, используются как руководящие формы для триаса.
Подкласс Ихтиозавры, или рыбоящеры. Ichthyopterygia. Подкласс объединил морских рептилий, утративших связь с сушей. Эти животные имели рыбообразное тело, мягкий спинной плавник и жесткие хвостовые плавники, парные конечности - ласты (рис. 34). Ласты – это видоизмененные конечности первых четвероногих. Длина тела некоторых форм достигала 23 м. Череп с длинной мордой и одной небольшой височной ямой (эвриапсидный тип). Зубы хорошо развиты, они конические, однорядные, лабиринтовые. Посредине хвост в той или иной степени изогнут вниз: у триасовых форм – незначительно, у юрских и меловых - очень резко. Чешуйчатый покров не установлен, но в коже была развита система роговых пластин. Кожа для лучшего скольжения в воде была, возможно, покрыта слизью.

Рис. 34. Ихтиозавр (реконструкция) [19]
У типичного ихтиозавра были очень большие глаза (до 20 см в диаметре), защищённые костяным кольцом, говорящим о том, что охотились они ночью. Ихтиозавры не имели жабр и дышали воздухом. Они не могли выходить на землю, поэтому перешли к живорождению, о чем свидетельствуют находки скелетов молодых особей внутри скелетов взрослых.
Ихтиозавры внешне похожи на рыб и дельфинов, к рыбам их первоначально и относили. Многие из ихтиозавров были хищниками, они питались моллюсками, в том числе белемнитами, рыбой, мелкими рептилиями, а также своими же сородичами.
Ихтиозавры известны с верхней части нижнего триаса, их расцвет приходится на раннюю юру, в позднем мелу они вымирают. Ихтиозавры, как и другие водные рептилии, имеют меньшую стратиграфическую ценность по сравнению с сухопутными рептилиями из-за их относительной малочисленности, тем не менее, они используются для расчленения юрских и меловых отложений.
Ихтиозавры захоронены в сланцах, мергелях, глинах, доломитах совместно с белемнитами, иглокожими, рыбами и др. Многочисленны как находки отдельных костей этих животных, так и целых скелетов, иногда с отпечатками мягких тканей. Скелеты могут быть заключены в известковые конкреции. Основные местонахождения остатков образовались на месте мелководных бухт.
Характерный представитель: ихтиозавр - Ichthyosaurus (MZ).
Подкласс Архозавры, илидревние ящерицы. Archosauria. Самая многочисленная группа рептилий с диапсидным типом (две височные впадины и часто предглазничное отверстие) черепа. По способу размножения архозавры были яйцекладущими животными. Архозавры произошли от чешуйчатых – эозухий в перми. В настоящее время они представлены крокодилами.
В составе подкласса выделяются надотряды и отряды, в том числе вымершие надотряды текодонтов, динозавров и птерозавров.
Надотряд Текодонты. Thecodontia – сборная группа, включающая первых архозавров, живших в перми и триасе. В последнее время ее представителей включают в группу круротарзов.
Внешне текодонты напоминали ящериц, динозавров и крокодилов. Передвигались на четырех или двух ногах. Они имели длинный узкий череп и текодонтные зубы. Это зубы, которые расположены в особых ячейках по краям челюстей - теках, которые снижали риск потерять их в процессе питания. Передние конечности у текодонтов были короче, чем задние.
Текодонты разделяются на два отряда: фитозавров (рис. 35) и псевдозухий (рис. 36). Фитозавры – крупные (до 5-6 м) крокодилообразные текодонты с удлиненным рылом и кожным панцирем, жившие в пресноводных бассейнах триаса и питавшиеся рыбой. Псевдозухии (рис. 36) - некрупные подвижные наземные животные ящерицеобразной и крокодилообразной формы с панцирем или без него. Среди последних присутствовали хищные, плотоядные и насекомоядные рептилии. Жили с перми по триас.
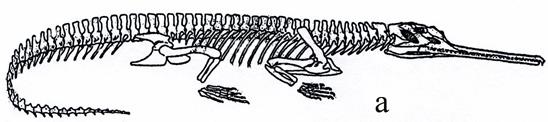

Рис. 35. Фитозавры: а - скелет [6], б – реконструкция [35]
Текодонты дали начало всем остальным группам архозавров: разнообразным динозаврам, летающим ящерам и крокодилам.
Представители надотряда используются в стратиграфии в основном триасовых отложений.
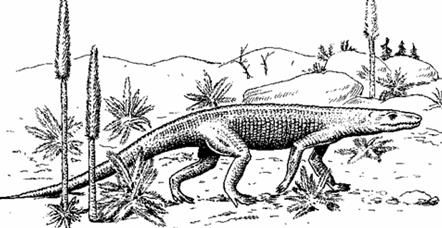
Рис. 36. Псевдозухия (реконструкция) [15]
Надотряд динозавров. Dinosauria (страшная ящерица). Это – самая многочисленная, господствующая в мезозое, группа рептилий. Жили с позднего триаса по поздний мел. Объединяет разнообразных по морфологии, размерам и образу жизни животных. Здесь есть небольшие рептилии величиной с кошку, рептилии средних размеров (большинство) и крупные - до 20-35 м (редко) в длину, двуногие и четвероногие, хищники и питавшиеся растениями, несшие костные панцири, «голые» и оперенные.
Выделяются два отряда динозавров: ящеротазовые и птицетазовые.
Отряд Ящеротазовые динозавры. Делятся на две группы: звероногие (тероподы) и ящероногие (завроподы). Появились в триасе, расцвет группы приходится на юру и ранний мел, в конце мела вымирают.
Т е р о п о д ы объединили двуногих животных с укороченными передними конечностями, которые передвигались на задних лапах, опираясь на сильный хвост. Среди них были хищники и растительноядные. Размеры достигали 14 м в длину. Некоторые тероподы имели перьевой покров.
Характерный представитель группы – тираннозавр (рис. 37). Известные в настоящее время его мелкие предки (например, дилонг) были оперены тонкими волосовидными перьями. У самого тираннозавра перьев могло и не быть (известные отпечатки кожи бедра тираннозавра несут типичный для динозавров рисунок из полигональных чешуй).

Рис. 37. Звероногий динозавр (теропод) – тираннозавр (Tirannosaurus, K2), реконструкция внешнего вида [24]
Большинство ученых с недавних пор стали относить к тероподам современных птиц (в качестве одного из подотрядов). Среди терапод можно найти формы, покрытые многочисленными пупырышками, волокнистым пухом или обладающих настоящими перьями.
Первым динозавром, у которого была установлена прижизненная окраска перьев, является синозавроптерикс (Sinosauropteryx), живший в раннем мелу (рис. 38). Ученым удалось выявить в останках его перьевого покрова окаменелые меланосомы — клеточные органеллы, в которых находятся молекулы меланина. На основе полученных исследований был сделан вывод, что синозавроптерикс имел яркое оперение: хвост был окрашен оранжево-белыми полосами, верхняя часть туловища имела рыжий цвет, живот был светлым, а ноги темными. Длина динозавра около 1 м.

Рис. 38. Теропод синозавроптерикс (Sinosauropteryx, К1) [30]
З а у р о п о д ы (рис. 39, 40, 41, 42) относились к растительноядным организмам. Они передвигались на четырех лапах. Размеры их головного мозга были очень маленькими.

Рис. 39. Ящероногий динозавр – диплодок (Diplodocus, J3).

Рис. 40. Кладка яиц крупного растительноядного динозавра-зауропода мелового периода (фото Т.Ю.Дрозд).
Зауроподы были самыми крупными животными, жившие когда-либо на нашей планете. Так, диплодоки и им подобные динозавры имели длину тела до 36 м и вес до 80 тонн. Считается, что они вели наземный, сухопутный образ жизни (как слоны) и перемещались по зарослям и степям от водоема к водоему.
Характерные представители: диплодоки, брахиозавры. Они, при общем внешнем сходстве, отличаются многими признаками. В частности, у диплодоков задние конечности длиннее передних, а у брахиозавров, наоборот, - передние существенно длиннее задних. Кроме того, у брахиозавров гребень на голове.

Рис. 41. На переднем плане – группа диплодоков [17].

Рис. 42. Брахиозавр (Brachiosaurus) [37]
Отряд Птицетазовые динозавры. Все рептилии этого отряда являются растительноядными. Они имели разные размеры и морфологию, многие имели защитные приспособления: панцирь, рога, щиты и др. Появляются в начале мезозоя, а к концу мела вымирают. Выделяются четыре группы этих динозавров: птиценогие динозавры, стегозавры, панцирные и рогатые динозавры.
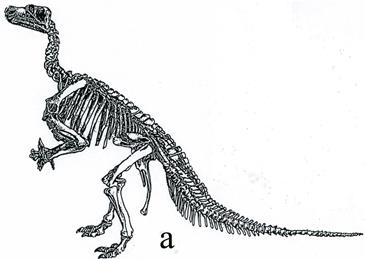
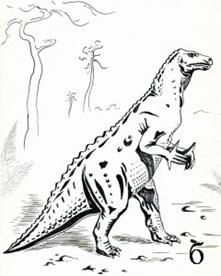
Птиценогие динозавры (игуанодонтиды) – высокие динозавры, ходившие на двух задних конечностях. Они жили на суше и в неглубоких водоемах. Один из их представителей, игуанодон (рис. 43), представлял собой крупное животное с длиной тела до 5-10 м и черепом похожим на лошадиный. Зубы напоминали ряд зубов млекопитающих. Передние конечности игуанодонов были короткими, пятипалыми, задние конечности были трехпалые с копытами. Обитал стадами вблизи водоемов. Ранний мел.
|
|
Рис. 43. Птиценогий динозавр игуанодон (Iguanodon, K1): а – скелет [6] [основы], б – реконструкция животного [31]
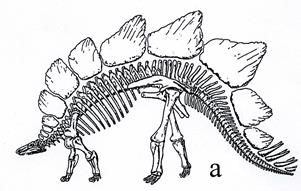
Стегозавры (рис. 44) первоначально передвигались на задних конечностях, а позже перешли к четвероногому хождению. Они имели большое туловище, маленькую голову и крупный хвост. Длина тела до 6 м.
| 
|
Рис. 44. Птицетазовый динозавр - стегозавр (Stegosaurus, J3):
а- скелет [6], б – реконструкция [42]
В качестве защитного приспособления на их спинах имелись костные пластинки размером до 1 м, а на конце хвоста – острые шипы. Распространение стегозавров – юра-ранний мел.
Панцирные динозавры (анкилозавры) (рис. 45) были крупными животными с широким уплощенным телом (длина до 5 м), покрытым массивным панцирем из костных пластин. Стратиграфическое распространение их узкое – мел.

Рис. 45. Панцирный динозавр – анкилозавр (Ankylosaurus, K2) [44]
В позднем мелу обитали рогатые динозавры (цератопсы) – крупные (до 7 м в длину) грузные рептилии, ходившие на четырех ногах. Характерная их особенность – наличие на черепе костного «воротника», который защищал область шеи. На конце морды и над глазами располагались рога. Типичный представитель группы – трицератопс (рис. 46).

| 
|
Рис. 46. Рогатый динозавр трицератопс (Triceratops, K2): а – скелет [6], б – реконструкция [41]
Надотряд Pterosauria. Птерозавры (крылатые ящеры) – летающие животные, внешне напоминающие птиц (рис. 47). Четырехпалые с сильно удлиненным четвертым пальцем передние конечности птерозавров были преобразованы в крылья. Между конечностями и боками тела была натянута летательная перепонка. Она представляла собой вытянутую складку кожи, которая крепилась на очень длинном четвертом пальце. Остальные три пальца короткие и оканчивались когтями. Птерозавры имели облегченные кости с воздушными полостями и хорошо развитую грудину с мощной мускулатурой. Челюсти – вытянутые, снабженные нередко зубами. Хвосты - разной длины.
Птерозавры являлись теплокровными рептилиями. У молодых особей имелся плотный «шерстистый» (волосовидный) покров.
По типу питания птерозавры насекомоядные, рыбоядные, некоторые питались трупами или планктоном, а также беспозвоночными.
Птерозавры произошли от текодонтов. Жили с позднего триаса до конца мела. Стратиграфическое значение их невелико.
Характерными представителями надотряда являются рамфоринхусы, птеродактили и птеранодоны. Первые (рис. 47а, 48) были размером с небольшую птицу, имели зубы и длинный хвост, заканчивающийся кисточкой. Летательные пальцы не складывались.
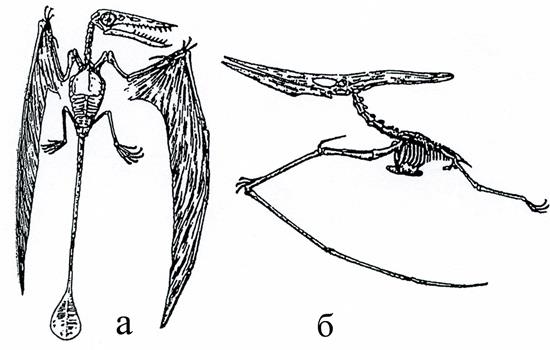
Рис. 47. Птерозавры, или крылатые ящеры:
а – длиннохвостый рамфоринхус (Rhamphorhinchus, J),
б – гигантский короткохвостый птеранодон (Pteranodon, K) [1]
Размеры птеродактилей - также небольшие, их клюв был снабжен зубами, но хвост был коротким или отсутствовал (рис. 50). Птеронодоны являлись беззубыми ящерами с коротким хвостом и высоким гребнем в верхней части черепа, имели большие размеры (размах крыльев достигал 16 м) (рис.47б, 49). У птеродактилей и птеранодонов летательные пальцы складывались.

Рис. 48. Рамфоринхус (Rhamphorhinchus, J) – реконструкция [16]

Рис.49. Птеранодон (Pteranodon, K) [28]

| 
|
Рис. 50. Птеродактиль (J): а - скелет, длина – 12 см [6], б – реконструкция внешнего вида [18]
Подкласс Crocodilia. Крокодилы. Т2-ныне. (рис. 51). Тело этих рептилий покрыто роговыми щитками, под которыми расположены костные пластинки. Туловище приплюснуто, хвост – длинный, задние конечности длиннее передних.
Крокодилы произошли от текодонтов в триасе, являются самыми организованными из современных пресмыкающихся. Сейчас населяют пресные и опресненные водоемы тропиков. Все крокодилы – хищники. Морские крокодилы используются в стратиграфии юры, пресноводные – верхнего мела.

Рис. 51. Телеозавры - ископаемые морские крокодилы
юрского периода [33]
2.3 Класс Aves. Птицы
Птицы в современной фауне представляют наиболее (после рыб) многочисленный класс позвоночных. Они хорошо приспособились к жизни в воздушной среде. Произошли от динозавров.
Основные признаки птиц и их отличия от пресмыкающихся:
- передние конечности птиц превращены в крылья, которые служат для летания и не приспособлены к другим функциям, которые обычны для передних конечностей других животных,
- теплокровны, поддерживают постоянную температуру тела,
- тело покрыто перьями, которые защищают от холода и помогают при полете,
- кровеносная, нервная системы и органы чувств более совершенны; сердце четырехкамерное,
- головной мозг больших размеров,
- скелет прочный и легкий, многие элементы скелета полые, содержащие в своих полостях воздух.
Часть из перечисленных признаков встречаются и у рептилий.
Остатки птиц находят редко, так как их кости быстро разрушаются. Иногда в ископаемом состоянии сохраняются перья или скорлупа.
Стратиграфическое значение ископаемых птиц невелико.
В классе выделено три подкласса: Довеерохвостые птицы (T3), Ящерохвостые птицы (J3-? K1) и Веерохвостые птицы (K-Q).
Довеерохвостые птицы (T3) представлены одним родом Protoavis (рис. 52), обнаруженом в позднем триасе. Эта птица была размером с сороку, имела длинный хвост, четырехпалые передние и задние конечности, позвонки как у птиц. Положение протоависа среди птиц спорно. Есть также мнение, что протоавис не птица, а представитель мелких хищных динозавров-теропод, от которого, как полагают, происходят настоящие, бесхвостые птицы

Рис. 52. Наиболее древняя птица протоавис –
Protoavis (реконструкция) [32]
Ящерохвостые птицы (J3). До находок протоависа эти птицы считались самыми древними. В своем строении они сочетали признаки пресмыкающихся и более поздних птиц. Характерный представитель - Archaeopteryx (рис. 53).

Рис. 53. Ящерохвостая птица археоптерикс (Archaeopteryx, J3):
а – общий вид (реконструкция), б – скелет [1]
Это птица величиной с ворону с длинным хвостом. Тело покрыто перьями. Череп маленький и плоский, что обычно для пресмыкающихся. Клюв не имел рогового чехла, зубы многочисленные. Передние конечности представляли собой крылья с тремя свободными пальцами с когтями. Задние конечности были четырехпалые. Археоптерисы вели как наземное, так и древесное существование. Возможно, они были способны только к планирующему полету.
Ящерохвостые птицы не являются прямыми предками настоящих птиц.
Веерохвостые (или новые) птицы. Это большая групп ископаемых и современных птиц. Среди них есть бегающие (страусы), плавающие (пингвины) и летающие (большинство) представители. Летающие веерохвостые птицы известны с позднего мела.
Среди ископаемых веерохвостых птиц встречались разнообразные птицы, в том числе с зубами. К зубастым птицам относят, например ихтиорнисов и гесперорнисов, живших в мелу. Ихтиорнисы (рис.53 а) – птицы до 25 см в длину, летающие, похожи на чаек. Гесперорнисы (рис. 53 б-в)– крупные до 180 см в длину, водоплавающие птицы, рыбоядные, похожи на гагар. У них был редуцирован плечевой пояс, исчезли кости крыла, крылья фактически отсутствовали. Ноги расходились в стороны под углом и работали как весла. Эта птица была приспособлена только к плаванию, способность к полету полностью утратила.

Рис. 54. Зубастые птицы: а – скелет ихтиорниса (Ichthyornis, K2),
б-в - гесперорнисы (Hesperornis, K2) (б – скелет, в – реконструкция) [1]
2.4 Класс Mammalia. Млекопитающие
Млекопитающие представляют собой класс животных, наиболее характерный для кайнозоя. Характерной особенностью их является живорождение (за исключением клоачных) и вскармливание детенышей молоком через специальные молочные железы. Первые находки млекопитающих известны из отложений верхнего триаса (рис. 55), в юре и мелу они еще немногочисленны. Произошли от звероподобных рептилий – териодонтов.

Рис. 55. Одно из первых млекопитающих – мегазостродон
(Megazostrodon, T) [27]
Млекопитающие приспособились к различным внешним условиям. Облик их разнообразен. Различны и способы питания, среди них есть растительноядные, плотоядные и всеядные. Прекрасно развиты дыхательная, пищеварительная, кровеносная, выделительная и нервная системы. Конечности пятипалые, но у некоторых животных в связи с образом жизни они видоизменяются. Зубы дифференцированы на резцы, клыки и щечные. Внешние покровы разнообразные: ногти, когти, копыта, роговые чешуи, рога. Тело голое или покрыто волосами.
Млекопитающие (кроме первозверей) - живородящие, теплокровные. У них развита забота о потомстве, детеныши вскармливаются молоком матери.
В составе класса выделяют шесть подклассов, три из которых, появившиеся с триаса, примитивные по строению, объединены в группу архаичных млекопитающих. Они имели небольшие размеры (как мыши), в конце мезозоя вымерли. Остальные три подкласса (первозвери, сумчатые, плацентарные) появились в мелу и дожили до наших дней.
Первозвери - примитивные млекопитающие, имеющие некоторые признаки рептилий. Произошли от архаичных млекопитающих. Размножаются яйцами, однако вскармливают детенышей молоком (точнее, молокообразной жидкостью. Яйцо помещают в сумку, где вылупившийся детеныш не всасывает, а слизывает молоко, выделяемое молочными железами, протоки которых открываются на коже брюшка (сосков нет). У некоторых первозверей есть зубы (молодые утконосы). Длина тела небольшая до 80 см. Образ жизни – наземный и полуводный. Современные представители: утконос, ехидна и проехидна.
Сумчатые, или низшие звери произошли от архаичных млекопитающих. Все сумчатые рождают живых детенышей и выкармливают их молоком. Детеныш заползает в сумку, где плотно присасывается к соску. По сравнению с высшими млекопитающими у сумчатых много древних, примитивных черт строения, которые резко отличают их от остальных зверей. Современные представители: кенгуру, коалы и др. Самый древний представитель сумчатых изображен на рис. 56.
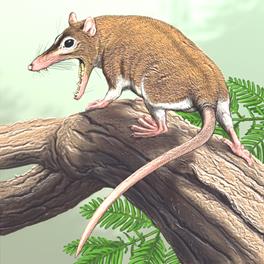
Рис. 56. Древнейший представитель сумчатых – синодельфис
(Sinodelphys, K) [29]
Плацентарные, или Высшие звери произошли от сумчатых. Для них характерно наличие плаценты, благодаря такому приспособлению детеныши рождаются достаточно развитыми и способными сосать молоко. Плацента выполняет различные функции: через нее эмбрион получает от матери питательные вещества и антитела, а также избавляется от продуктов жизнедеятельности. Для плацентарных характерно высокое развитие мозга.

Рис. 57. Первое плацентарное животное юрамайя (Juramaia, J2) [36]
В подклассе выделяется 27 отрядов, из них 10 – вымершие. Наиболее примитивные плацентарные – отряд насекомоядные, они явились исходной группой, от которой произошли другие отряды плацентарных. Самыми распространенными представителями насекомоядных является семейство ежовых.
Среди плацентарных, в частности, имеются следующие отряды: Грызуны (рис. 58а) (зайцы), Хищные (рис. 58б) (гиены, кошки), Китообразные (дельфины), Древние копытные, Непарнокопытные (рис. 59, 62б) (носороги, лошади), Парнокопытные (рис. 60а, 61) (олени, верблюды), Хоботные (рис. 60б, в) (мамонт), Приматы (рис. 64) (обезьяны, люди).

Рис. 58. Высшие звери: а – зайцеобразный грызун эуримилус (Eurymylus, палеоген), б – гиеновидный хищник гиениктитерий (Нуaenictitherium, N) [9]

Рис. 59. Высшие звери. Отряд непарнокопытные: гигантский носорог индрикотерий (Indricotherium, поздний палеоген):
а – скелет [7], б – реконструкция [1]

Рис. 60. Парнокопытные и хоботные: а - отряд Парнокопытные,
большерогий олень мегалоцерас (Megaloceras, Q1) [1], б-в - отряд Хоботные: б - мамонт (Mammutus, Q2) [1], в – мастодон (Mastodon, N-Q2) [7]
Для определения млекопитающих используют различный костный материал, но особенно большое значение имеют зубы (рис. 61, 62). Поэтому при сборе остатков этих животных необходимо обратить особое внимание на эти элементы скелета. Зубы мелких млекопитающих выявляют путем промывания и просеивания осадка, в котором они найдены или их наличие предполагается.

Рис. 61. Зубы мерикоидодона (Merycoidodon), представителя надотряда
Копытные (отряд Парнокопытные), жившего в палеогене-начале неогена

| 
|
Рис. 62. Зубы разных млекопитающих:
а – глиптогона (Glyptogon, Q, вымерший) из семейства броненосцев; б – лошади (Equus, палеоген-ныне) из отряда непарнокопытных
Отряд Приматы. Это один из наиболее прогрессивных отрядов плацентарных млекопитающих, включающий полуобезъян, обезьян, гоминид, в том числе человека, и несколько вымерших групп, близких к полуобезъянам. Для приматов характерны пятипалые конечности, выполняющие хватательные движения, при этом большой палец кисти противопоставлен остальным (у многих), пальцы снабжены ногтями, развитые большие полушарии мозга, бинокулярное зрение, волосяной покров, недоразвитое обоняние.
Приматы возникли от общего с шерстокрылами (летающие лемуры) предка в верхнемеловое время.
Первое известное приматоподобное млекопитающее – пургаториус (Purgatorius) (рис. 63) – найдено в отложениях верхнего мела. Это был небольшой, размером с мышь, зверёк, живший на деревьях и питавшийся насекомыми. Пургаториус является возможным предком приматов. Более того, представителей этого рода иногда называют первыми " примитивными приматами".

Рис. 63. Пургаториус (Purgatorius) – наш далекий предок [21]
Человек принадлежит к семейству людей, в составе которого имеется и Homo sapiens L. – человек разумный. Представителей людей делят на древнейших (питекантропов), древних (палеоантропов или неандертальцов) и современных (неоантропов). Существуют разные классификации гоминид. Люди известны с верхней части неогена (плиоцен).
Большинство исследователей считает древнейшим предком человека ардипитека (рис. 64), который жил в раннем плиоцене примерно5, 8—4, 4 млн лет назад. Доказательством этому служит сходство зубов ардипитека с зубами австралопитека. Кроме того, в стопе ардипитека имеется добавочная малая берцовая кость (os peroneum), которая присутствует в стопе человека, но отсутствует в стопе современных человекообразных обезьян.
Ардипитеки жили в тропических лесах, были способны к прямохождению (однако способны были лазить и по деревьям) и питались фруктами и орехами. Они рассматриваются предками не только человека, но и шимпанзе. Их череп занимает промежуточное положение между этими группами гоминид. Человек по множеству признаков более похож на ардипитека, чем шимпанзе. 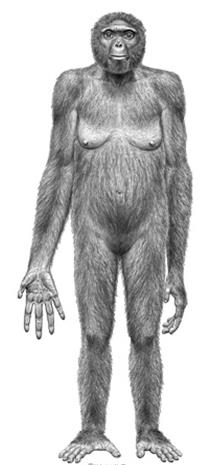
Полные скелеты млекопитающих находят редко. Преимущественно встречаются такие разрозненные элементы скелета, как челюсти, зубы, рога, ключицы, конечности, части черепа, позвонки, ребра. Нередко на месте необходимо применение специальных методов препарирования, чтобы предупредить разрушение скелетных остатков при высыхании.
Остатки млекопитающих находят в различных породах. Так, довольно часто они встречаются в ледниковых отложениях (например, во флювиогляциальных отложениях террас), в торфах, в отложениях пещер, в вечной мерзлоте и т.д.
Рис. 64. Ардипитек Арди [10]
Млекопитающие широко используются для расчленения кайнозоя, особенно, четвертичного периода. Это объясняется тем, что остатки их в кайнозое многочисленны, а скорость эволюции очень высокая. Например, в позднем кайнозое (с позднего олигоцена по голоцен) Европы выделено 24 зоны млекопитающих, каждая из которых имеет свой характерный набор животных.
ЗАКЛЮЧЕНИЕ
В органическом мире позвоночные занимают особое место. Морфологические особенности их уникальны и по сравнению с беспозвоночными они являются более совершенными представителями животных.
Количество позвоночных в целом невелико (всего 45 тыс. современных видов), но они весьма разнообразны по приспособительным и жизненным формам. Позвоночные играют важную роль в биосферных процессах, т.к. обычно завершают трофические связи в биоценозах. Позвоночные – важнейший фактор географической среды. Они имеют меньшее стратиграфическое значение по сравнению с беспозвоночными, тем не менее, используются для расчленения, корреляции и определения возраста отложений палеозоя, мезозоя и кайнозоя.
ЛИТЕРАТУРА И ИНФОРМАЦИОННЫЕ ИСТОЧНИКИ
1. Друшиц В.В., Обручева О.П. Палеонтология. – М.: МГУ, 1971. – 413 с.
2. Еськов К.Ю. История Земли и жизни на ней: учебное пособие. – М.: МИРОС, 2000. – 349 с
3. Лебедев О.А. Первые четвероногие: поиски и находки. Природа, № 11, 1985, с. 26-36.
4. Михайлова И.А., Бондаренко О.Б., Обручева О.П. Общая палеонтология. – М.: МГУ, 1989. – 384 с.
5. Основы палеонтологии. Бесчелюстные. Рыбы / под ред. Д.В.Обручева. – М.: Наука, 1964. – 522 с.
6. Основы палеонтологии. Земноводные, пресмыкающиеся и птицы / под ред. Ю.А.Орлова. – М.: Наука, 1964. – 722 с.
7. Основы палеонтологии. Млекопитающие / под ред. В.И.Громова. – М.: Наука, 1962. – 421 с.
8. Парфенова М.Д. Историческая геология с основами палеонтологии: учебное пособие. – Томск: Изд-во ТПУ, 1998. – 550 с.
9. Шпанский А.В. Основы палеонтологии тетрапод: учебное пособие. – Томск: Изд-во Томск. гос. ун-та систем упр. и радиоэлектроники, 2005. – 216 с.
10. https://antropogenez.ru/zveno-single/57
11. https://batrachos.com/node/132
12. https://batrachos.com/node/401
13. https://bioword.narod.ru/B/B062.htm
14. https://bse.sci-lib.com/article089781.html
15. https://bugabooks.com/book/296-yevolyuciya-zhizni/71-obnovlenie-flory-i-fauny-v-triase.html
16. https://dinoblog.org/ptero/ramforinh.html
17. https://dinosaurs.about.com/od/dinosaurbasics/tp/10-Facts-About-20-Dinosaurs.htm
18. https://dinozavry.tcoa.ru/yurskii_period/pterodaktil.html
19. https://en.wikipedia.org/wiki/Ichthyosaurus
20. https://forum.zoologist.ru/viewtopic.php? pid=23819
21. https://hronoblog.blogspot.com/2011/01/1455-655-15.html
22. https://kottus.wordpress.com/2007/06/08/389/
23. https://my-paleo-world.ucoz.com/publ/8-1-0-56
24. https://paleontology.narod.ru/dino/rex.htm
25. https://ru.wikipedia.org/wiki
26. https://ru.wikipedia.org/wiki/Tiktaalik_roseae
27.https://ru.wikipedia.org/wiki/%D0%A4%D0%B0%D0%B9%D0%BB: Megazostrodon.jpg
28. https://samtec.su/? bdf=pteranodon-picture
29. https://scienceblogs.com
30. https://scienceblog.ru/page/138/
31. https://shkolazhizni.ru/archive/0/n-38906/
32. https://squamata.ru/dyno/227-protoavis.html
33. https://squamata.ru/vympres/101-vymkro.html
34. https://superspeak.ru/lofiversion/index.php/t635.html
35. https://vadim-proskurin.livejournal.com/366371.html
36. https://zhelezyaka.com/news.php? id=5732
37. https://www.ancientbeasts.ru/photo/3-0-4
38. https://www.answersingenesis.org/articles/tj/v17/n2/tetrapod
39. https://www.answers.com/topic/prolacertiformes
40. https://www.denizhaber.com.tr/dis-basindan/21957/pliosaur-jurassic-sahili-deniz-canavari.html
41. https://www.dinozavro.ru/melovoy/triceratops.php
42. https://www.evolbiol.ru/_pnaug3.htm
43. https://www.juraparkbaltow.pl/index.php? go=dino& d=mastodonsaurus
44. https://www.librero.ru/article/myth/dinoxavr/pic/
45. https://www.mathematical.com/dinodviniaprima.html
46. https://www.physiologus.de/pikaia.htm
47. https://www.uadream.com/tourism/europe/Russia/element.php? ID=47490
48. https://www.zoomet.ru/ixt/ixtiolog_24.html
49. https://900igr.net/kartinki/biologija/Uslozhnenie-zhivotnykh-v-protsesse-evoljutsii/050-Kisteperye-ryby.html
ОГЛАВЛЕНИЕ
| ВВЕДЕНИЕ………………………………………………………………… 1 Раздел Anamnia. Низшие позвоночные…………………………………. 1.1 Инфратип Agnata. Бесчелюстные………………………………. 1.2 Инфратип Gnatostomata. Челюстноротые……………………… 1.2.1 Надкласс Pisces. Рыбы………………………………….. 1.2.1.1 Класс Placodermi. Пластинокожие……………. 1.2.1.2 Класс Acanthodei. Акантоды………………….. 1.2.1.3 Класс Chondrichthyes. Хрящевые……………... 1.2.1.4 Класс Osteichthyes. Костные………………….. 1.2.2 Надкласс Tetrapoda. Четвероногие……………………. 1.2.2.1 Класс Amphibia. Земноводные………………… 2. Раздел Amniota. Высшие позвоночные………………………………… 2.1 Класс Reptilia. Пресмыкающиеся…………………. 2.2 Класс Aves. Птицы…………………………………. 2.3 Класс Mammalia. Млекопитающие……………….. ЗАКЛЮЧЕНИЕ…………………………………………………………….. ЛИТЕРАТУРА………………………………………………………………. |
Учебное издание
|
|