Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Розтягання поверхневих клітин ектодерми.
|
|
Перебудова багатошарової стінки даху бластули приводить до початку епіболії (мал. 7).

Рис. 7. Інтеркаляція декількох глибоких шарів клітин (Б, В - по Гілберту, 1993):
А – схема процесу; Б – будова даху бластули жаби на стадії середньої бластули;
В – теж на стадії середньої гаструли
У ході реаранжировки клітини зовнішнього ряду стають більш плоскими, стінка бластули стає тоншою, а клітинний шар зміщається вбік бластопору, який формується.
3. Конвергенція клітин крайової зони бластули. Спостерігається в ході гаструляції в амфібій (мал. 8).
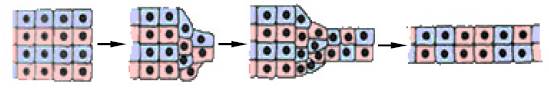
Рис. 8. Конвергентне розтягання (схема)
Являє собою сходження до центра та подовження інволюючої ділянки бластули, розташованої безпосередньо над областю бластопору. У результаті стягування клітин у більш вузьку смужку відбувається її розтягання в передньому напрямку.
4. Поляризація клітин. Будь-яка активна зміна форми епітеліального шару в період гаструляції та нейруляції починається з того, що його клітини поляризуються, тобто витягаються в перпендикулярному або косому напрямку відносно до поверхні шару. Поляризація клітин заснована на складних перебудовах цитоскелету та клітинної мембрани: збиранню мікротрубочок і мікрофіламентів та їхньої орієнтації по довгій осі клітини, яка поляризується, а також рухами, так званих, інтегральних білків. В результаті цих рухів відбувається перерозподіл іонних каналів і насосів у плазмалемі. Перші концентруються на зовнішній стороні клітини, а другі на бічних і базальних сторонах. Поляризація обумовлює протікання в клітинах спрямованого ендо- і екзоцитозу.
Поляризація зачіпає не одну клітину, а цілий клітинний шар, тобто поляризація однієї клітини спонукує сусідню до такого ж перетворення. Цей процес здійснюється лише при наявності клітинних контактів і тому має назву контактної клітинної поляризації.
Клітини не можуть залишатися в стані поляризації необмежено довго. Через певний час відбувається деформація поляризованої клітини, що зменшує співвідношення її поверхні до обсягу (скорочення). Цей процес здійснюється скорочувальним апаратом клітини - мікрофіламентами. У ході послідовних скорочень шар вигинається, відбувається елементарне морфогенетичне переміщення.
5. Скорочення поляризованих клітин. У результаті скорочення апікальних поверхонь поляризованих клітин відбувається зміна форми всього клітинного шару, утвореними ними (мал. 9).

Рис. 9. Згортання шару клітин (схема):
А, Б – послідовні стадії процесу
Подібні скорочення клітин нейроектодерми відіграють важливу роль у формуванні нервової трубки. У цьому випадку рушійною силою є скорочення кільця актинових мікрофіламентів, що оточують апікальні краї клітин і пов'язаних з мембраною клітини за допомогою інтегрального білка спектрина.
6. Утворення колбоподібних клітин. У ході поляризації формуються так називані колбоподібні клітини (мал. 10).
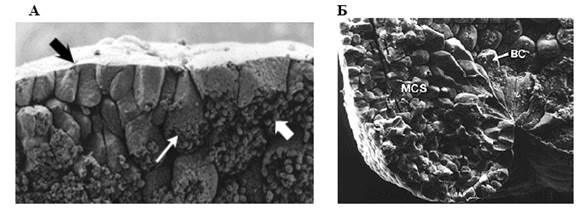
Рис. 10. Колбоподібні клітини (за Гілбертом, 1993):
А – бокалоподібні клітини (тонка світла стрілка) до початку інвагінації поверхневого шару (темна стрілка) до середини глибоких шарів (товста світла стрілка) бластули;
Б –бласопор зародка амфібій, що формується (ВС – бокалоподібні клітини, МС – мезодермальні клітини)
Основна частина тіла кожної клітини зміщена до середини зародка, зберігаючи контакт із поверхнею за допомогою вузького цитоплазматичного тяжа. Вони здатні скорочуватися, активно переміщатися досередини, і тягти за собою інші клітини шару. У результаті починає формуватися та поступово збільшуватися порожнина архентерону.
Проте виявилося, що після того, як сформувалися колбоподібні клітини, вони не мають особливого відношення до збільшення розмірів гастроцелю. Часткове або повне видалення цих клітин не запобігає інволюції прилеглих до них клітин у бластопор.
Головним фактором руху клітин до середини зародка є інволюція глибоких клітин крайової зони бластули. Ці клітини укручуються досередини та мігрують до анімального полюса по внутрішніх поверхнях глибоких клітин, які ще не інволювали (мал. 11). Видалення лідируючих глибоких клітин крайової зони зупиняє формування архентерона.
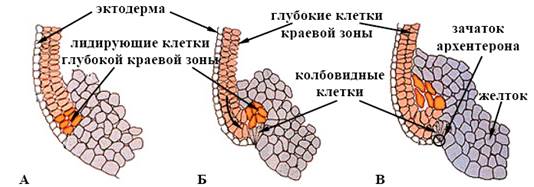
Рис. 11. Модель клітинних рухів у період гаструляції (за Гілбертом, 1993):
А, Б, В – послідовні стадії процесу
Таким чином, колбоподібні клітини відповідальні лише за утворення початкового поглиблення бластопора, а рушійна сила інволюції укладена в глибокому шарі крайових клітин.
7. Здатність клітин до амебоїдних рухів. Адгезивність і рухливість неоднакові в клітин різних листків. Так клітини ектодерми, контактуючи один з одним, утворюють безперервний шар, тобто епітелізуються.
Вони здатні поширюватися над мезодермою та ентодермою. Клітини мезодерми мають тенденцію інвагінувати у будь-яке скупчення клітин, що знаходиться поблизу, а клітини ентодерми відносно нерухомі. Для гаструляції характерна активність особливих груп клітин. Так у морських їжаків на початку гаструляції надзвичайно активні клітини первинної мезенхіми (мал. 12).
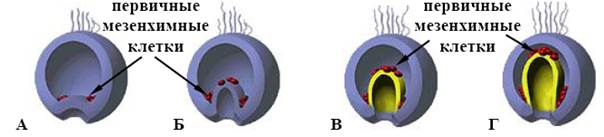
Рис. 12. Поведінка мезенхімних клітин при гаструляції в морського їжака (схема):
А – первинні мезенхімні клітини мігрували досередини бластоцеля; Б – вторинні мезенхімні клітини попадають досередини разом з інвагінуючою первинною кишкою;
В, Г – мезенхімні клітини зберігають своє положення на вершині архентерону, а скорочення їхніх нитковидних відростків, прикріплених до внутрішньої поверхні ектодерми, сприяє інвагінації архентерону.
Вони переміщаються від внутрішньої стінки зародка в області бластопору в бластоцель, випускають ламеллоподії (мал. 13) і активно мігрують до того місця, де вони утворюють кістяк. Слідом за ними починає активно інвагінувати інша частина зародка.

Рис. 13. Мігруючі за допомогою ламеллоподій лідируючі клітини мезодерми зародка шпорцевої жаби
8. Роль позаклітинного матриксу. Потрапивши досередини бластоцеля, клітини за допомогою ламеллоподій мігрують по позаклітинному матриксу. Для їхньої успішної міграції важливі 2 білки.
Один з них – фібронектин. Це високомолекулярний глікопротеїн (400 кДа), що є звичайним компонентом базальних мембран.
Дослідження показали, що під час гаструляції спорідненість мігруючих клітин до даного білка різко зростає, а сам процес міграції залежить від концентрації фібронектину (мал. 14).
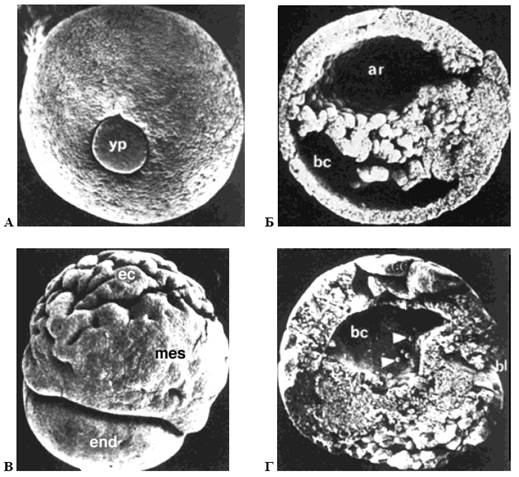
Рис. 14. Роль фібронектинового матриксу в процесах гаструляції в амфібій (за Гілбертом, 1993):
А, Б – нормально розвинена бластула (вид з боку жовточної пробки та поперечний розріз, yp - жовточна пробка, ar – архентерон, bc - бластоцель); В, Г – теж після ін'єкції синтетичного пептиду, що перешкоджає зв'язуванню мігруючих клітин з фібронектином (bl – бластопор, ес – ектодерма, mes – мезодерма, end – ентодерма)
Клітини зв'язуються з певною ділянкою фібронектина, що містить специфічну послідовність із 10 амінокислот.
Друга важлива складова частина позаклітинного матриксу – сульфатовані глікопротеїни клітинної поверхні мігруючих клітин.
Важливу роль у міграції клітин досередини бластоцеля, особливо у вищих хребетних (птахів і ссавців) грають позаклітинні складні полісахариди. Одним з них є гіалуронова кислота, лінійний полімер глюкуронової кислоти та N-ацетілглюкозаміну. Вона накопичується в бластоцелі, покриваючи суцільним шаром поверхню клітин, що вступають у бластоцель, підтримуючи їх у диспергованому стані за рахунок здатності значно збільшувати свій обсяг у воді.
Як ні різноманітні типи гаструляції, існують загальні зміни на клітинному рівні, які приводять до морфогенетичних переміщень. Клітинні поділи, що виникають у певному регіоні у зв'язку з особливими зовнішніми умовами і впливами, створюють механічні напруги в шарі. Ці напруги ведуть до виникнення поляризації, яка несе інформацію про напрямок майбутнього переміщення, а його реалізація, тобто властиво переміщення, відбувається під час скорочення. Морфогенетичні рухи протягом усього розвитку є активними. Їхнє джерела енергії та виконавчих механізмів перебувають усередині тієї ділянки, що зазнає тієї або іншої деформації.
Факторами організації можуть бути механічні натягання тканин зародка. Першим фактором, що обумовив натягання ембріональних тканин, є тургорний тиск у порожнині бластоцеля, що розтягує дах бластоцеля. Якщо послабити його, виникають порушення в морфогенезі та структурі органів.
Таким чином, в основі більшості формоутворюючих процесів лежать клітинні розподіли, що викликають, у свою чергу, виникнення механічних напруг у шарі клітин, що приводять до зміни форми зародка.
У регуляції шляхів розвитку важливу роль грають кілька десятків білків, іменованих факторами росту, які в організмі тварин регулюють клітинні поділи та диференціровку, а також виникнення тканин. Подібно нейромедіаторам у нервовій системі такі фактори, очевидно, використовуються в різних обставинах для передачі різних сигналів між клітинами.
|
|