Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Сравнительная характеристика ДНК-полимераз E. coli
|
|
| ДНК-полимераза | |||
| I | II | III | |
| Структурный ген* | polA | polB | polC (dnaE) |
| Число различных видов субъединиц | ≥ 10 | ||
| Молекулярная масса | 103.000 | 88.000† | 791.500 |
| 3/®5/-экзонуклеазная корректирующая активность | + | + | + |
| 5/®3/-экзонуклеазная активность | + | – | – |
| Скорость полимеризации (нуклеотидов/сек) | 16-20 | 250-1000 | |
| Эффективность (процессивность) (количество добавленных нуклеотидов до отсоединения полимеразы) | 3-200 | 1.500 | ≥ 500.000 |
* для ферментов включающих более одной субъединицы, приведенные в таблице обозначения генов относятся только к полимеризующей активности. Обратите внимание на то, что обозначение dnaE является несколько устаревшим обозначением принятого в настоящее время символа polC.
† молекулярная масса указана только для полимеризующей субъединицы. ДНК-полимераза II имеет несколько общих с ДНК-полимеразой III субъединиц, включая: b, g, d, d/, c и y-субъединицы.
Таким образом, ДНК-полимераза I не является основополагающим ферментом репликации, вместо этого он выполняет массу «подчищающих» функций, принимая участие в процессах репликации, рекомбинации и репарации. Более того, специфические функции данного фермента расширяются благодаря его специфической 5/®3/-экзонуклеазной активности. Данная активность в отличие от корректирующей 3/®5/-экзонуклеазной активности связана с отдельным структурным доменом, который может быть отщеплен от интактного фермента посредством мягкого протеолиза. После отщепления 5/®3/-экзонуклеазного домена образующийся так называемый большой фрагмент ДНК-полимеразы I или фрагмент Кленова (мол. масса 68.000 Да) сохраняет полимеразную и корректирующую 3/®5/-экзонуклеазную активности. Специфическая 5/®3/-экзонуклеазная активность интактной ДНК-полимеразы I ответственна за замещение небольших сегментов ДНК (РНК) спаренных с матричной цепью в процессе так называемой nick-трансляции. Характерно, что другие ДНК-полимеразы утратили 5/®3/-экзонуклеазную активность.
Основной фермент репликации ДНК у E. coli – ДНК-полимераза III является гораздо более сложно устроенным, чем ДНК-полимераза I комплексом, включающим 10 разных видов (типов) субъединиц (табл. В) (рис. 1). Полимеризующая и корректирующая активности этого фермента связаны с его a- и e- субъединицами, соответственно. Субъединица q взаимодействуя с a- и e- субъединицами образует кор-фермент, который способен катализировать реакцию полимеризации, но с ограниченной эффективностью. Два кор-фермента могут быть соединены с другим набором субъединиц: g-комплексом, состоящим из семи субъединиц шести различных типов – t 2 gdd/cy, который способствует значительному увеличению процессивности полимеразы. (Мерой процессивности ДНК-полимеразы является длина фрагмента вновь синтезированной макромолекулы, которую фермент способен образовать в одном цикле, не диссоциируя от матрицы). За процесс сборки холофермента и его димеризацию отвечают t-субъединицы. Субъединицы t и g кодируются одним и тем же геном, однако g-субъединица является укороченным вариантом субъединицы t. Таким образом t-субъединица включает домен идентичный g и дополнительный сегмент ответственный за связывание с кор-ферментом. Полный ансамбль из 13 белковых субъединиц девяти (9) различных видов обозначают ДНК-полимеразой III*.
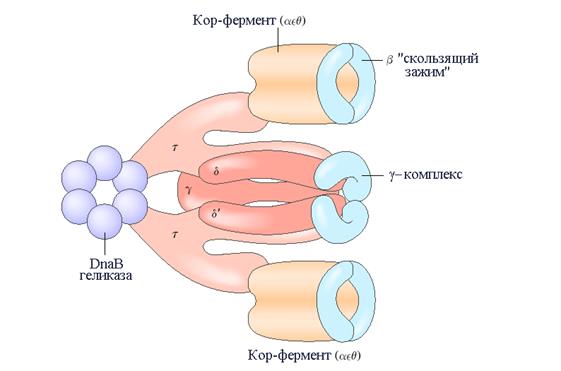
| Рис. 1 | Строение ДНК-полимеразы III E. coli. Показано, что холофермент ДНК-полимеразы III включает два домена, представленных кор-ферментами состоящими из субъединиц a, e и q. Два кор-фермента связаны в димер с помощью g-комплекса построенного из семи субъединиц шести различных типов – t2gdd/cy (субъединицы c и y не показаны). Субъединицы t и g кодируются одним и тем же геном, однако g-субъединица является укороченным вариантом субъединицы t. Таким образом t-субъединица включает домен идентичный g и дополнительный сегмент ответственный за связывание с кор-ферментом. С каждым кор-субансамблем взаимодействует так называемый b-«скользящий зажим» состоящий из двух b-субъединиц. В целом холофермент ДНК-полимеразы III связан с dnaB-геликазой с помощью субъединицы t. PDB ID 2POL |
ДНК-полимераза III* способна полимеризовать ДНК, но с намного более низкой процессивностью, чем необходимо для удвоения всей хромосомы E. coli.
Таблица В
Характеристика субъединиц ДНК-полимеразы III E. coli
| Субъединица | Число субъединиц в холоферменте | Молекулярная масса субъединицы | Ген | Функции субъединиц | |
| a | 129.900 | polC (dnaE) | полимеразная активность | кор-фермент | |
| e | 27.500 | dnaQ (mutD) | 3/→ 5/-экзонуклеазная корректирующая активность | ||
| q | 8.600 | holE | |||
| t | 71.100 | dnaX | стабилизация связывания с матрицей, димеризация кор-фермента | g-комплекс, который обеспечивает взаимодействие b-субъединиц вокруг отстающей цепи в каждом фрагменте Оказаки | |
| g | 47.500 | dnaX* | формирование «скользящего зажима» | ||
| d | 38.700 | holA | открывает «скользящий зажим» | ||
| d/ | 36.900 | holB | формирование «скользящего зажима» | ||
| c | 16.600 | holC | взаимодействие с SSB-белками | ||
| y | 15.200 | holD | взаимодействие с g и c | ||
| b | 40.600 | dnaN | «скользящий зажим» ДНК для оптимальной процессивности |
* g-субъединица кодируется частью гена субъединицы t так, что 66% N-концевой последовательности t совпадает с полной аминокислотной последовательностью g. g-субъединица образуется путем сдвига рамки считывания, что приводит к ранней терминации трансляции.
Увеличение процессивности фермента достигается путем присоединения 4-х b-субъединиц на каждый холоэнзим ДНК-полимеразы III. b-субъединицы взаимодействуют попарно с образованием кольцеобразной структуры, которая обхватывает ДНК и действует на подобие «хомута». Каждый димер взаимодействует с субансамблем кор-фермента ДНК-полимеразы III* и «скользит» вдоль ДНК в ходе репликации. «Скользящий зажим» препятствует отделению полимеразы от молекулы ДНК, существенно увеличивая тем самым процессивность вплоть до 500.000 добавленных нуклеотидов.
|
|