Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Механизм сопряжения дыхания и фосфорилирования в митохондриях 8 страница
|
|
. Обтурационная желтуха (обтурация — в переводе закупорка) возникает в результате нарушения оттока желчи в кишечник. Она сопровождается выходом конъюгированного билирубина из клеток печени обратно в кровь. В крови повышено содержание конъюгированного билирубина, который кай хорошо растворимое соединение выделяется в больших количествах с мочой Из-за этого моча имеет цвет пива с ярко-желтой пеной. Кал, в котором отсутствуют желчные пигменты, приобретает серовато-белый, глинистый цвет.
Желтуха новорожденных' считается физиологической. Она возникает вследствие возрастного недостатка фермента конъюгации билирубина — глюку ронозилтраи сфер азы. Поэтому повыщспие распада эритроцитов, вызванное любыми причинами, ведет к повышению неконъюгнрованного билирубина в крови и желтухе. Физиологическая желтуха новорожденных обычно проходит через 2 недели по мере увеличения количества глюкуронозилтранс- феразы в печени. У недоношенных детей она продолжается дольше. Длительное повышение неконъюгированного билирубина в крови может1 быть опасным вследствие токсического действия билирубина на развивающийся мозг. Иногда у детей появляются судороги или необратимые расстройства нервной системы. У взрослых клетки мозга малопроницаемы для билирубина н, как правило, осложнений при гипербилирубинемии не происходит. Чтобы повысить активность глкжуронозилтрансферазы, иногда прибегают к введению фенобарбитала, ' который увеличивает количество фермента и уменьшает проявления желтухи.
Нарушение пигментного обмена наблюдается при дисбактериозе кишечника, возникающего в- результате подавления его нормальной микрофлоры (например, при длительном лечении антибиотиками тетрацикяинового ряда) При дисбактериозе выделяются с калом промежуточные продукты восстановления билирубина, а в случае глубокого подавления микрофлоры сам билирубин, который окисляется в биливердин, окрашивающий кал в зеленоватый цвет.
ГЛАВА 21. ОБМЕН ПУРИНОВЫХ И ПИРИМИДИНОВЫХ МОНОНУКЛЕОТНДОВ
Для млекопитающих не требуется поступления с пищей азотистых оснований или нуклеотидов, хотя при приеме пищи, богатой нуклеиновыми кислотами, они всасываются в готовом виде. В их тканях расход пуриновых и пиримиди- новых нуклеотидов непрерывно возобновляется путем синтеза из простых фрагментов.
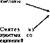
Нуклеозидфосфаты, содержание которых в нормальных клетках невелико и относительно постоянно, являются связующим звеном на путях поступления и использования мононуклеотидов:
Нуклеиновые Синтез нуклеиновых
|
| Нуклеозидфосфаты — |
1. Биосинтез мононуклеотидов
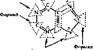
Биосинтез пуриновых мононуклеотидов. Первоначальным соединением синтезе пуриновых нуклеотидов служит фосфорибозилпирофосфат (ФРПФ), образующийся из рибозо-5-фосфата и АТФ. Первым пуриновым мононуклеотидом, завершающим длинную цепь реакций синтеза, является инозинмонофосфат (ИМФ), из которого затем образуются остальные пуриновые нуклеозидфос- фаты через АМФ и КМФ (ксавтдзинмонофосфат). Происхождение атомов луринового кольца показано на рис. 62.
Биосинтез пиримидиновых мононуклеотидов. Первоначальными соединениями в биосинтезе пиримидичовы* млночуклептипов являются карбамоил- фосфат и аспарагиновая кислота. Из них образуются через длинную цепь реакций УМФ и остальные пиримидиновые мононуклеотиды (рис. 63). Источниками всех атомов пиримид«нового кольца являются карбамоилфосфат и аспарагиновая кислота.
Биосинтез деэоксирибонуклеотидов, являющихся предшественниками ДНК, происходит путем восстановления рибозы до 2'-дезоксирибозы уже в составе готового нуклеотнда. Восстановителем служит особый белок — тио- редоксин, содержащий в молекуле две^ЭН-группы.
2. Распад нуклеиновых кислот и нуклеотидов
Нуклеиновые кислоты гидролизуются в тканях организма с помощью нукле- аз — деэоксирибонуклеаз (ДНКазы) и рибонуклеаз (РНКаэы). Эти ферменты катализируют расщепление внутренних или концевых межнуклеотидных связей (3', S'-фосфодиэфнрных) в ДНК и РНК, Иуклеазы, расщепляющие связи внутри полинуклеотидной цепи, называются андокуклеозами, а отщепляющие концевые нуклеотиды — зкзонуклеазами. Некоторые экзочуклеазы способны парализовать как ДНК. так и РНК. Под действием нуклеаз поли- куклеотидная цепь распадается до олигонуклеотидов и мононуклеотидов (нувдеозмд-б'- или 3'-монофосфатов).
Мононуклеотиды распадаются до конечных продуктов обмена -гидролити-
|
| у / |
| Глицнн |
| Аспартат |
ФРПФ
ИМф
|
| I |
АМФ КМФ
 АДФ АТФ ГМФ
АДФ АТФ ГМФ
|
Рис, 62. Схема синтеза пуриновых соиоиуклеотидов (о) и источники атомов пуркноеого кольца (& \ нуклеотидов
Карбамоклфосфат Аспарагиновая кислота
Дигидрооротовая кислота
' 1
Оротрвая кислота»
Оротидин-5^ф осфат I t
• тмф «■» умф «■ ■ ■» амф
/\ /\ /\
ттф «. ТДф удф «■ утф цдф < —».цтф
Рнс. 63. Схема синтеза пиримндиновых нуклеотидов
ческнм или фосфоролнтическим путями (в первом случае для разрыва связей используется вода, а во втором — фосфорная кислота),
Мононуклеотиды расщепляются с помощью нуклвотидаз {5'-нуклеоти- дазы и. З'-нуклеотидааы) и нуклеозид гидролаз до свободных оснований, пентозы и фосфата,
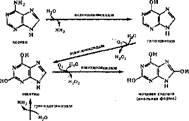
|
| гуанин |
| Она является конечным продуктом обмена руриновых нуклеотидов в организме человека и выделяется с мочой, Превращение пирнмндиновых основа- |
Пуриновые основания окисляются до мочевой кислоты:
ний идет до (3-аланина, С02 и аммиака. 0-Аланин используется для синтеза дипептидов мышц — карноз'ина и ансерина — или выделяется с мочой.
Таким образом, содержание мочевой кислоты в крови и моче отражает интенсивность расщепления нуклеиновых кислот в организме. Если принимать специальную беспуриновую днету, то по выделению с мочой мочевой кислоты можно судить об обмене эндогенных пуринов. Мочевая кислота — плохо растворимое в воде соединение, поэтому нормальные концентрации ее в жидких средах организма приближаются к пределу растворимости. Повышение - ее содержания в крови — гиперурикемия — вызывает отложение в виде кристаллов урата натрия в тканях, особенно при местном снижении рН среды. Заболевание, сопровождающееся отложением уратов в суставах, поч- кйл, называется ноии£ рой. Оно широко распространено, особенно у мужчин. Иногда к подагре приводит врожденная, частичная или полная недостаточность фермента, участвующего в повторном использовании свободных оснований в синтезе нуклеотидов. Препараты, снижающие образование мочевой кислоты при окислении пуринов (например, аллопуринол) или стимулирующие выведение ее почками (например, антуран, цинхофен), облегчают течение подагры.
ГЛАВА 22. ВЗАИМОСВЯЗЬ ОСНОВНЫХ ПУТЕЙ ОБМЕНА ВЕЩЕСТВ В ОРГАНИЗМЕ
Обмен важнейших структурных мономеров живых систем, т. е. аминокислот, моносахаридов (глюкозы), жирных-кислот, мононуклеотидов, тесно взаимосвязан. Эта взаимосвязь осуществляется через так называемые ключевые метаболиты, которые служат общим звеном на пЗтях распада или.синтеза мономеров (рис. 64). К таким метаболитам можно отнести пируват, аце- тил-КоА, о-глицеролфосфат и промежуточные продукты цикла Кребса (оксалоацетат, малат, фумарат, сукцинил-КоА, 2-оксоглутарат, изоцитрат и цитрат). Пируват является точкой пересечения путей распада и синтеза глюкозы (и других моносахаридов) и некоторых аминокислот. Более разветвленный узел метаболических связей представляет собой ацетил-КоА. Через него перекидывается мостик от моносахаридов и аминокислот к липидам, т. е. открывается путь превращения аминокислот (а следовательно, и белков) и глюкозы (т. е. вообще углеводов) в липиды. Как бы вспомогательным связующим звеном между углеводами и липидамн служит а-глице- ролфосфат. Через него происходит превращение углеводов в некоторые липиды |(триацилглицерины, фосфоглицериды) и, наоборот, липидов, содержащих глицерин, в углеводы.
Обширные возможности для взаимопревращения одних мономеров в другие предоставляют промежуточные продукты цикла Кребса. Через цикл Кребса сообщаются все основные пути распада и синтеза веществ. Поэтому согласно схеме, приведенной на рис. 64, можно, например, представить, что жирные кислоты, являющиеся источником ацетил-КоА, превращаются в углеводы (через оксалоацетат цикла Кребса происходит новообразование глюкозы), в аминокислоты (через оксалоацетат и 2-оксоглутарат в аспара- гнновую и глутаминовую кислоты), в порфирины (через сукцинкл-КоА).
Мономеры являются структурными звеньями биополимеров, поэтому.возл,
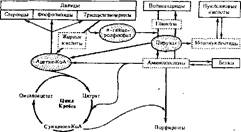 Рис. 64. Упрошенная схема взаимосвязи основных путей обмена веществ
Рис. 64. Упрошенная схема взаимосвязи основных путей обмена веществ
|
можна относительная взаимозаменяемость белков, " углеводов и липидов как главных компонентов пищи. Границы взаимозаменяемости позволяют понять не только возможные нарушения обмена при однообразном питании, но и механизм компенсации обмена веществ при голодании, т. е. образования одних веществ за счет эндогенных резервов других. Каковы же границы взаимозаменяемости? Эти рамки ограничивают прежде всего аминокислоты. Восемь незаменимых аминокислот не могут синтезироваться в организме млекопитающих из других веществ, да и синтез полузаменимых аминокислот ограничен. Следовательно, без этих аминокислот или без содержащих нх пищевых белков организму ие обойтись. Очевидно, аминокислоты (прежде всего незаменимые) являются первичными по отношению к остальным биомолекулам, и с их.возникновения, возможно, началась эволюция живых систем.
Из схемы взаимосвязи путей обмена становится понятным, что если потребляются в полном ассортименте и достаточном количестве аминокислоты (точнее пищевые белкн) и углеводы, то организм - может обходиться достаточно долгое время без липидов (за исключением полиненасыщенных липидов). Аминокислоты и углеводы служат источником ацетил-КоА, а углеводы — к тому же источником и НАДФ • Н2, что позволяет синтезировать липиды в нужном количестве. Та же схема показывает, почему организму- человека вообще не требуется поступления с пищей пуриновых и пиримидй- новых нуклеотидов (структурных мономеров нуклеиновых кислот), ибо при наличии в пище белков и углеводов они синтезируются в необходимых количествах. Следовательно, белки и углеводы позволяют обеспечивать длительное время образование жизненно важных'Компонентов тканей организма и его нормальную деятельность. Конечно, это возможно, если с белками я углевозе] дами поступают и такие компоненты пиши, как витамины, вода и неорганические ионы. Человек может долгое время обходиться и без углеводов, если поступают а необходимом количестве остальные компоненты пиши.
Г. ПЕРЕНОС ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ И ЕГО ЗНАЧЕНИЕ ДЛЯ ЖИЗНЕДЕЯТЕЛЬНОСТИ КЛЕТОК
Способность к переносу генетической информации, т. е. к передаче наследственных свойств, является уникальным свойством живых организмов. Хра- нение и передача генетической информации, которую можно рассматривать как разновидность биологической памяти, возложена природой на нуклеиновые кислоты.
Генетическая программа, заложенная в ДНК хромосом ядра и митохондрий (или хлоропластов) клеток организмов, одинакова. Их Специализация определяется различиями в реализации этой генетической программы в ходе развития клеток. Поэтому зрелые, дифференцированные клетки, например клетки нервной ткани, печени, отличаются друг от друга набором молекулярных компонентов, формирующих характерный структурный образ данной клетки, особенностями метаболизма н в конечном счете специфическими функциями, Формирование этих особенностей разных клеток можно упрощенно выразить в виде схемы (см, схему 4),
Схема 4. Роль генетическое информации в формировании специальных структур клетки
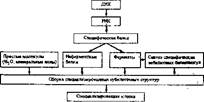
|
Из нее видно, насколько важным является процесс переноса генетической информации для развития и нормальная деятельности клеток opra. киэма.
ГЛАВА 23. ПЕРЕНОС ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ И БИОСИНТЕЗ БЕЛКА В КЛЕТКАХ
I. Виды переноса генетической информации
Можно отметить три варианта переноса генетической информации, обнаруженные в различных организмах.
.1. Перенос генетической информации в пределах одного класса нуклеиновых кислот, т. е. от ДНК к ДНК или у некоторых вирусов от РНК к РНК. называется репликацией или самоудвуеннем, Иногда такой вид переноса наследственной информации образно называют.«переносом из хранилища в хранилище», Подразумевая под этим, что ДНК (или РНК у некоторых вирусов) выполняет функцию хранилища генетической информации. Репликация происходит во время деления клеток (в 5; фазу митотического цикла) и размножения вирусов, когда необходимо целиком передать информацию от одного организма к другому. Возможна репликация отдельных участков ДНК, называемая амплификацией.
2. Перенос информации между разными классами нуклеиновых кислот: ДНК—РНК, называется транскрипцией или переписыванием. В отличие от репликации при транскрипции происходит копирование не целиком всей ин-
| t |
opMauHH, заложенной в молекуле ДНК, а только ее отдельных участков, ходе транскрипции образуются все типы РНК: главные (мРНК, тРНК, рРНК) и минорные.
Из' этого следует, что циЬтроны ДНк содержат информацию не только о структуре полипептидной цепи, но и о структуре тРНК( рРНК и минорных типов РНК.
. Транскрипция бывает прямая (от ДНК к РНК) и обратная (от РНК к ДНК)- Впервые обратная транскрипция была выявлена у РНКовых опухоле- ролных вирусов (называемых онкорнавнрусами), которые встраиваются в ДНК клетки-хозяина путем обратной транскрипции. Чужеродный участок ДНК'- копия вирусной £ Нк— вызывает опухолевуй трансформацию клетки, Возможно, обратная транскрипция имеет значение не только при опухолевой трансформации клеток, & о и при их нормальной жизнедеятельности -или в ходе их дифференцировки. В- принципе обратная транскрипция возможна при использовании любого типа РНК, а не только мРНК.
3. Перенос генетической информации от мРНК к белку, т. е. в пределах разных классов макромолекул, называется трансляцией или переводом. При трансляции происходит как бы перевод информации с «языка» нуклеиновых кислот, - имеющего нуклео'тидный алфавит, на «язык» полипептидной цепи, в котором использован аминокислотный алфавит. Транслируется только мРНК. Остальные типы РНК являются готовыми продуктами генов, образующимися при транскрипции. рРНК и тРНК играют вспомогательную роль при трансляции, трансляция, по термодинамическим и биологическим соображениям, может быть толыр прямой — от мРНК к белку. Обратная трансляция запрещена.
Перенос генетической информации в клетках — процесс непрерывный, и его можно выразить следующей схемой:
дик -J
гРНК -и РНК |)РНК
I
Подобное направление переноса генетической информации от ДНК через РНК к белку называется цептралопош iшстулшим молекулярной генетики, который был сформулирован Криком. Согласно ему не может быть переноса информации от белка к РНК, ' но допускается перенос от РНК к ДНК. Если вдруг обнаружится перенос информации от белка к РНК, то придется пересмотреть весь фундамент молекулярной генетики.
Итак, первичными продуктами действия генов являются два типа макромолекул. Это прежде всего белок и некоторые типы РНК: рРНК, тРНК и минорные РНК.
Все виды передачи генетической информации основаны на матричном механизме. Это означает, что для каждого ил них необходима матрица. При репликации матрицей служит одна из цепей ДНК (или РНК у вируеив), при транскрипции — участок ДНК (прямая транскрипция) или РНК (обратная транскрипция), а при трансляции — мРНК, т. е. матрицей может быть только нуклеиновая кислота. Матрица позволяет с большой точностью (что очень важно, поскольку речь идет о наследственных свойствах) и экономичностью (что не менее важно) воспроизводить имеющуюся в клетке генетическую информацию. Точность копирования соответствующей нуклеиновой матрицы обеспечивает правило комплементарности азотистых оснований нуклеотидов, согласно которому происходит спаривание А с Т (или с У в РНК) -и Г с Ц. Благодаря этому порядок чередования нуклеотидов в каждой новой полину- клеотидной цепи комплементарен матрице.
Представим себе, что не было" бы матричного механизма переноса информации. В этом случае даже для сборки, например, одной цепи специфической ДНК бактерии, содержащей около миллиона нуклеотидов, нужно иметь полмиллиона высоко специфичных молекул ферментов (ибо каждая фосфоди- эфирная связь между рядом стоящими нуклеотидами в полннуклеотндной цепи должна образовываться строго специфично). Следовательно, клетка должна иметь невероятно большой избыток ферментов только для репликации, а сам процесс был бы труден и длителен, к тому же высокая точность его вряд ли была бы возможна.
2. Молекулярные основы репликации
Теоретически возможны несколько вариантов репликации ДНК: консервативная, когда дочерняя двойная спираль ДНК образуемся бел разделения цепей родительской ДНК; полуконсервативная> при которой цепи родительской ДНК расходятся, и на каждой из родительских цепей образуются комплементарные цепи дочерней ДНК, и дисперсивная, которая предусматривает расщепление в нескольких местах цепей родительской ДНК и образование на ней новых цепей ДНК.
В 1957 г. Меселсон и Сталь установили, что в живых организмах репликация ДНК происходит по полуконсервативному механизму. Сначала он представлялся просторна расплетающихся цепях ДНК, которые являются матрицами, образуются комплементарные новые цепи ДНК- Нуклеотиды при этом выстраиваются по матрице соответственно правилу комплементарности, а «сшиваются» друг с другом фосфодиэфирнымй связями с помощью специального фермента ДНК-полимеразы. Впоследствии оказалось, что ДНК-полиме- раза не способна начать синтез новой ДНК из свободных нуклеотидов; она лишь способна удлинять полинуклеотидную цепь, т. е. для нее нужна затравка. В настоящее время репликация ДНК представляется сложным, многоступенчатым процессом, который имеет отличительные признаки у про-. кариотов и эукариотов.
Для репликации ДНК необходим ряд условий;
1) наличие дезоксирибонуклеозидтрнфосфатов (дАТФ, дТТФ, дГТФ и дЦ'ГФ) как структурного материала для сборки новых цепей ДНК;
2) расплетение двойной спирали ДНК;
3) образование затравки;
4) наличие ферментов, участвующих в образовании затравки и синтезе новых полинуклеотидных цепей ДНК.
Механизм репликации ДНК у прокариотов. Каждая стадия процесса репликации протекает с участием соответствующих ферментов.
1. Расплетающие белки разрывают водородные связи между комплементарными основаниями двойной спирали ДНК- В результате двойная спираль расплетается и расходится на отдельные цепи. {Внешне это напоминает расстегивание застежки «молния».) Расплетенный участок ДНК называется репликативной вилкой. В его образовании участвует сразу до 200 молекул расплетающего белка, поэтому каждая ветвь репликативной вилки, на которой происходит начало синтеза новой ДНК, имеет протяженность до 2000 не- спаренных оснований. Механизм действия расплетающих белков детально не изучен. Возможно, для расплетения ДНК используется энергия АТФ.
2. *Затравочная» ДНК-зависимая РНК-полимераза — особый вариант РНК-полимеразы {обычно эти ферменты участвуют в транскрипции), которая образует небольшую РНК-затравку («праймер»), комплементарную участку ДНК в репликативной внлке. Синтез РНК-затравки идет от конца 5' к концу 3'. Порядок чередования нуклеотидов в РНК задается ДНК-матрицей, а сшивка их 5'-> -3'-фосфодиэфйрными связями осуществляется РНК-полиме- разой.
3. ДНК-Полимеразы. Известны три формы ДНК-полимераз у прокариотов: I, II и III. Все они обладают двумя видами активности: полимеразной и нуклеазной. Полимеразная состоит в образовании фосфодиэфирных связей 5'-»-3' между дезоксирибонуклеотидами, нуклеазная — в гидролизе фосфодиэфирных связей.
ДНК-полимераза I расщепляет РНК-затравку при репликации {действует как РНКаза) и на ее месте синтезирует комгьлементарный фрагмент ДНК. ДНК-полимераза - II имеет очень низкую полимеразную активность. Функция ее в репликации неизвестна.
: ДНК-Полимераза III является основным ферментом репликации, синте- зирун на разошедшихся цепях ДНК комплементарные участки новой ДНК в направлении б'-^З'.
4. Рибонукледаа Н. Участвует в гидролизе РНК-затравки в ходе репликации наряду с ДНК-полимераэой I.
5. ДНК-Лигаэы {соединяющие ферменты). Обнаружено несколько ферментов, функция которых состоит в соединении друг с другом новосинте- зированных 'фрагментов ДНК- ДНК-Лигазы образуют фосфодиэфирные связи 3'-*-5', используя НАД* в качестве источника адеиилила,
Общая схема репликации представляется следующим образом (рис, 65). Под действием расплетающих белков образуются сразу в нескольких местах молекулы ДНК (как правило, на внутренних участках) реплнкативные вилки. Начинается репликация (инициация) с образования РНК-затравки с по* мощью РНК-полнмеразы в направлении б'^З'. После завершения синтеза короткой цепи РНК на матрице ДНК фермент отделяется от ДНК, Далее происходит присоединение к РНК-затравке дезоксирибонуклеотидов с помощью ДНК-полимеразы III в направлении 5'-*-3', Образуется гибридная цепь: РНК —ДНК, Причем ДНК-полимераза III синтезирует короткие фрагменты ДНК (фрагменты Оказаки) на другой родительской цепи репликатив- ной вилки. Интересно, что по ходу синтеза ДНК-полимераза III может исправлять ошибки при неправильном спаривании нуклеотидов. Если произошла ошибка, то этот нуклеотид тут же отщепляется ферментом благодаря нуклеазной активности, а при правильном спаривании" нового нуклеотида присоединяет его к уже имеющемуся фрагменту ДНК.
РНК-Затравка после действия ДНК-полимеразы III полностью удаляется или специфической рибонуклеазой Н, или ДНК-полимеразой I, На месте бывшей РНК-затравки наращивается цепь ДНК с помощью ДИК-полиме- разы I. Соединение синтезированных фрагментов ДНК (фрагментов Оказаки) н направлении происходит с йомощью ДНК-лигазы,
Точность репликации очень высока. Частота спонтанных ошибок при репликации составляет около Ю~в—10~8, Это означает, что возможно ошибочное включение только одного нуклеотида на каждый новосинтезированный фрагмент ДНК, содержащий 10е—10s нуклеотидов.
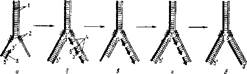 Рис. 65, Схема репликации ДНК; и — расхождение ueneft родительской ДНК и синтез РНК-'затраекк; б — образование комплементарной гибридной цепи РНК—ДНК и синтез фрагментов Окзэакн в направлении $'~*-3'\ в —гидролиз РНК-затравки; г — достраивание ком племен тарной цепи ДНК на месте РНК-затравки, д сшивание фрагментов комплементарной цепи ДНК; I — родительские цепн; 3 — расплетающий белок; 8— РНК; 4 — фрагменты Оказаки
Рис. 65, Схема репликации ДНК; и — расхождение ueneft родительской ДНК и синтез РНК-'затраекк; б — образование комплементарной гибридной цепи РНК—ДНК и синтез фрагментов Окзэакн в направлении $'~*-3'\ в —гидролиз РНК-затравки; г — достраивание ком племен тарной цепи ДНК на месте РНК-затравки, д сшивание фрагментов комплементарной цепи ДНК; I — родительские цепн; 3 — расплетающий белок; 8— РНК; 4 — фрагменты Оказаки
|
Репликация ДНК у эукариот, Репликация ДНК в хромосомах и митохондриях эукариотов происходит тоже полуконсервативным способом. Имеются некоторые особенности репликации ДНК ядер и митохондрий {или хлоропластов) эукариот. В клетках млекопитающих обнаружены те же ферменты репликации ДНК {расплетающие белки,. РНК-полнмераза, ДНК-полимеразы, рибонуклеаэа Н, ДНК-лигазы), Однако эти ферменты отличаются то молекулярной структуре и свойствам от ферментов прокариотов. Так, ДНК-полнме- раэы ядер {а их тоже обнаружено три — а, р и у) и митохондрий (у-типа) клеток млекопитающих не обладают нуклеазной активностью. Наиболее активна у них ДНК-полимераза о, Она, видимо, выполняет ту же функцию при репликации, что и ДНК-полимераза Ш прокариотов.
Репарация ДНК- Репарацию, или исправление поврежденных участков одной из цепей ДНК, можно рассматривать как. ограниченную репликацию. Наиболее изучен процесс репарации при повреждении цепи ДНК ультрафиолетовым излучением, например, эпителиальных клеток кожи. При УФ облучении образуются «сшивки» между соседними остатками тнмина в ДНК, Эти димеры тимина уже не могут выполнять функцию матрицы при репликации. Поэтому в клетках имеются ферменты, исправляющие поврежденные участки ДНК. Сначала такой участок удаляется ДНКазамн. Затем фермент типа ДНК-полимеразы I заполняет пробел путем синтеза участка ДНК в направлении На последнем этапе сшиваются концы ДНК-лигазой.
3. Молекулярные основы транскрипции
В процессе жизнедеятельности зрелой клетки только часть генетической информации, записанной в ДНК хроматина, реализуется при транскрипции в виде копий РНК- Участки неактивной, или репрессированной, ДНК входят в состав глобулярных нуклеосом хроматина, а активной — в состав межнук- леосомных фрагментов или «развернутых» линейных нуклеосом.
Элементарную единицу транскрипции у прокариотов и эукариотов, т. е. отрезок ДНК, подвергающийся транскрипции, принято называть транскриптоном. Иногда транскриптоны прокариотов называют также оперонами. Длина транскриптонов колеблется от 300 до 10® нуклеотидов {последняя цифра характерна для высших эукариотов, у которых, как правило, размеры транскриптонов намного больше, чем у прокариотов).
Отдельные участки транскриптонов несут разную функцию. Одна группа участков относится к информативным, а другая — к неинформативным. К информативным относятся структурные цистроны или гены, несущие информацию о структуре полипептндной цепи или нематричных РНК {рРНК, тРНК, других низкрмолекулярных стабильных РНК); неинформативные выполняют другие функции и не содержат генетической информации. Особенно велика неинформативная часть транскриптона у высших эукариотов. Совершенно неожиданно оказалось, что структурные гены в транскриптоне могут быть двух типов: непрерывными и «разорванными», или прерывистыми. Непрерывность генов считалась абсолютным признаком. Действительно, трудно было представить, что, например, полипептидная цепь записана в соответствующем цнстроне не вся, а кусками. Тем не менее во многих структурных генах, особенно эукариотов, генетическая информация записана с перерывами. Участки в структурных генах, несущие информацию, называются экзонами.
а неинформативные — интронами Возможно, нитроны играют дополнительную регуляторную функцию для экзонов.
В ДНК хромосом обнаружены подвижные фрагменты, которые названы лабильными генами или трансппзонами. Выявлено несколько разновидностей таких генов, отличающихся нуклеотидным составом и длиной полинуклеотид- ной цепи. Миграцию транспозонов объясняют механизмом обратной транскрипции, т. е. сначала образуется транскрипт мобильного гена, а затем он используется как матрица для встраивания ДНК-копии а другом участке хромосом. Функция «прыгающих» генов еще не ясна Известно, однако, что они могут вызывать перестройки в геноме, изменять функцию тех участков ДНК. рядом с которыми они встраиваются. В частности, вклиниваясь рядом с онкогеном (участком ДНК, вызывающим перестройку нормальной клетки в опухолевую), мобильные гены активируют их функцию, что может привести к опухолевому перерождению ткани Вместо с тем мобильные гены, участвуя в перестройке отдельных фрагментов хромосом, влияют на изменчивость организмов и их эволюцию.
Схема функциональной организации транскриптона у прокариотов н эу- кариотов представлена на рис, 66. Начальный участок транскриптона, с которого начинается транскрипция, называется промотор. К нему присоединяются белки, облегчающие начало транскрипции, и фермент транскрипции РНК-полнмераза. Оператор — участок ДНК, связывающий белки-регуляторы транскрипции. Таким бел ком-регулятором транскрипции у прокариотов является репрессор.
У эукариотов после промотора находится участок транскриптона, который называют акцепторной или управляющей зоной. С ней взаимодействуют различные регуляторы, влияющие на транскрипцию. В акцепторной зоне
Промотор Акцепторная Структурные rt
И i Э; И! Э I И: Э: И | Э
Прокариоты
«-, -.,.... ■.. -------------..,.,,.. Транекрнптон
(оперон)
Промотор Оператор Структурные п
Рис. 66. Функциональная организация транскриптонов у прокариотов к эукариотов (И — интрон; Э — экзон)
имеется фрагмент ДНК (его называют усилитель или «энхансер»), который облегчает транскрипцию с участием РНЙ-полимеразы.
К оператору, или акцепторной зоне, примыкают структурные цистроны, или гены, содержащие перемежающиеся участки интронов и экзонов. В одном транскриптоне может быть один структурный цистрон (моноцистронный транскриптон) или несколько (полицистронный транскриптон). В конце транскриптона имеется последовательность нуклеотидов, которая является своего рода сигналом об окончании транскрипции, — терминатор. РНК, образующаяся при транскрипции, называется транскриптом. Транскрипт — комплементарная копия транскриптона от промотора до терминатора.
|
|