Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Рефлексы Геринга — Брейера.
|
|
Раздувание легких у наркотизированного животного рефлекторно тормозит вдох и вызывает выдох. Нервные окончания, расположенные в бронхиальных мышцах, играют роль рецепторов растяжения легких. Их относят к медленно адаптирующимся рецепторам растяжения легких, которые иннервируются ми-елинизированными волокнами блуждающего нерва.
Рефлекс Геринга — Брейера контролирует глубину и частоту дыхания. У человека он имеет физиологическое значение при дыхательных объемах свыше 1 л (например, при физической нагрузке). У бодрствующего взрослого человека кратковременная двусторонняя блокада блуждающих нервов с помощью местной анестезии не влияет ни на глубину, ни на частоту дыхания.
У новорожденных рефлекс Геринга — Брейера четко проявляется только в первые 3—4 дня после рождения.
Проприоцептивный контроль дыхания. Рецепторы суставов грудной клетки посылают импульсы в кору больших полушарий и являются единственным источником информации о движениях грудной клетки и дыхательных объемах.
Межреберные мышцы, в меньшей степени диафрагма, содержат большое количество мышечных веретен. Активность этих рецепторов проявляется при пассивном растяжении мышц, изометрическом сокращении и изолированном сокращении интрафузальных мышечных волокон. Рецепторы посылают сигналы в соответствующие сегменты спинного мозга. Недостаточное укорочение инспираторных или экспираторных мышц усиливает импульсацию от мышечных веретен, которые через у-мотонейроны повышают активность о-мотонейронов и дозируют таким образом мышечное усилие.
Хеморефлексы дыхания. Рог и Рсог в артериальной крови человека и животных поддерживается на достаточно стабильном уровне, несмотря на значительные изменения потребления Оз и выделение С02. Гипоксия и понижение рН крови (ацидоз) вызывают усиление вентиляции (гипервентиляция), а гипероксия и повышение рН крови (алкалоз) — понижение вентиляции (гиповентиляция) или апноэ. Контроль за нормальным содержанием во внутренней среде организма 02, СОг и рН осуществляется периферическими и центральными хеморецепторами.
Адекватным раздражителем для периферических хеморецепторов является уменьшение Ро; артериальной крови, в меньшей степени увеличение Рсо2 и рН, а для центральных хеморецепторов — увеличение концентрации Н* во внеклеточной жидкости мозга.
Артериальные (периферические) хеморецепто-ры. Периферические хеморецепторы находятся в каротидных и
аортальных тельцах. Сигналы от артериальных хеморецепторов по синокаротидным и аортальным нервам первоначально поступают к нейронам ядра одиночного пучка продолговатого мозга, а затем переключаются на нейроны дыхательного центра. Ответ периферических хеморецепторов на понижение Рао^ является очень быстрым, но нелинейным. При Рао; в пределах 80—60 мм рт. ст. (10, 6—8, 0 кПа) наблюдается слабое усиление вентиляции, а при Рао; ниже 50 мм рт. ст. (6, 7 кПа) возникает выраженная гипервентиляция.
Расо2 и рН крови только потенцируют эффект гипоксии на артериальные хеморецепторы и не являются адекватными раздражителями для этого типа хеморецепторов дыхания.
Реакция артериальных хеморецепторов и дыхания на гипоксию. Недостаток С> 2 в артериальной крови является основным раздражителем периферических хеморецепторов. Импульсная активность в афферентных волокнах синокаротидного нерва прекращается при Раод выше 400 мм рт. ст. (53, 2 кПа). При нормоксии частота разрядов синокаротидного нерва составляет 10% от их максимальной реакции, которая наблюдается при Раод около 50 мм рт. ст. и ниже-Гипоксическая реакция дыхания практически отсутствует у коренных жителей высокогорья и исчезает примерно через 5 лет у жителей равнин после начала их апаптации к высокогорью (3500 м и выше).
Центральные хеморецепторы. Окончательно не установлено местоположение центральных хеморецепторов. Исследователи считают, что такие хеморецепторы находятся в ростральных отделах продолговатого мозга вблизи его вентральной поверхности, а также в различных зонах дорсального дыхательного ядра.
Наличие центральных хеморецепторов доказывается достаточно просто: после перерезки синокаротидных и аортальных нервов у подопытных животных исчезает чувствительность дыхательного центра к гипоксии, но полностью сохраняется реакция дыхания на гиперкапнию и ацидоз. Перерезка ствола мозга непосредственно выше продолговатого мозга не влияет на характер этой реакции.
Адекватным раздражителем для центральных хеморецепторов является изменение концентрации Н4 во внеклеточной жидкости мозга. Функцию регулятора пороговых сдвигов рН в области центральных хеморецепторов выполняют структуры гематоэнцефали-ческого барьера, который отделяет кровь от внеклеточной жидкости мозга. Через этот барьер осуществляется транспорт 02, С02 и Н^ между кровью и внеклеточной жидкостью мозга. Транспорт СОз и H+ из внутренней среды мозга в плазму крови через структуры гематоэнцефалического барьера регулируется с участием фермента ка рбоангидразы.
Реакция дыхания на COi. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз тормозят центральные хеморецепторы.
Для определения чувствительности центральных хеморецепторов к изменению рН внеклеточной жидкости мозга используют метод возвратного дыхания. Испытуемый дышит из замкнутой емкости, заполненной предварительно чистым Од. При дыхании в замкнутой
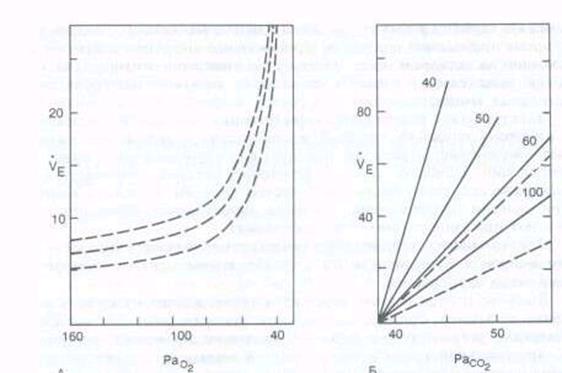
Рис. 8.12. Изменение вентиляции легких (ve. л'мин) в зависимости от парциального давления Од (А) и COz < Б) в альвеолярном воздухе при различном содержании Ог в альвеолярном воздухе (40, 50. 60 и 100 мм рт. ст.).
системе выдыхаемый СО; вызывает линейное увеличение концентрации СОа и одновременно повышает концентрацию Н* в крови, а также во внеклеточной жидкости мозга. Тест проводят в течение 4—5 мин под контролем содержания СО; в выдыхаемом воздухе-
На рис. 8.12 показано изменение объема вентиляции при различном уровне напряжения С02 в артериальной крови. При Расоа ниже 40 мм рт. ст. (5, 3 кПа) может возникнуть апноэ в результате гипокапнии. В этот период дыхательный центр мало чувствителен к гипоксической стимуляции периферических хеморецепторов.
8.6.3. Координация дыхания с другими функциями организма
В филогенетическом развитии организма человека и животных дыхательный центр приобретает сложные синаптические взаимоотношения с различными отделами ЦНС.
В отличие от других физиологических функций организма дыхание находится под контролем автономной (вегетативной) и соматической нервной системы, поэтому у человека и животных дыхание нередко называют вегето-сомати ческой функцией. Существует тесное взаимодействие регуляции дыхания гуморальной и рефлекторной природы и процессами сознательной деятельности мозга. Однако во время сна или в состояниях, связанных с отсутствием сознания у человека, сохраняется внешнее дыхание и обеспечивается нормальное поддержание газового гомеостаза внутренней среды. С другой стороны, человек имеет возможность по собственному желанию
изменять глубину и частоту дыхания или задерживать его, например во время пребывания под водой. Произвольное управление дыханием основано на корковом представительстве проприоцептивного анализатора дыхательных мышц и на наличии коркового контроля дыхательных мышц.
Электрическое раздражение коры больших полушарий у человека и животных показало, что возбуждение одних корковых зон вызывает увеличение, а раздражение других — уменьшение легочной вентиляции. Наиболее сильное угнетение дыхания возникает при электрической стимуляции лимбической системы переднего мозга. При участии центров терморегуляции гипоталамуса возникает ги-перпноэ при гипертермических состояниях.
Однако многие нейрофизиологические механизмы взаимодействия нейронов переднего мозга с дыхательным центром остаются пока мало изученными.
Дыхание опосредованно через газы крови влияет на кровообращение во многих органах. Важнейшим гуморальным, или метаболическим, регулятором локального мозгового кровотока являются Н* артериальной крови и межклеточной жидкости. В качестве метаболического регулятора тонуса сосудов мозга рассматривают также С02. В последнее время эта точка зрения подвергается сомнению, поскольку СО-1 как молекулярное соединение практически отсутствует во внутренней среде организма. Молекулярный С02 (0—С=-0) встречается в организме в альвеолярном воздухе, а в тканях только при переносе СОд через аэрогематический и гистогематический барьеры. В крови и межклеточной жидкости СО; находится в связанном состоянии, в виде гидрокарбонатов, поэтому правильнее говорить о метаболической регуляции Н^ тонуса гладких мышц артериальных сосудов и их просвета. В головном мозге повышение концентрации Н^ расширяет сосуды, а понижение концентрации I^ в артериальной крови или межклеточной жидкости, напротив, повышает тонус гладких мышц сосудистой стенки. Возникающие при этом изменения мозгового кровотока способствуют изменению градиента рН по обе стороны гематоэнцефалического барьера и создают благоприятные условия либо для вымывания из сосудов мозга крови с низким значением рН, либо для понижения рН крови в результате замедления кровотока.
Функциональное взаимодействие систем регуляции дыхания и кровообращения является предметом интенсивных физиологических исследований. Обе системы имеют общие рефлексогенные зоны в сосудах: аортальную и синокаротидные. Периферические хеморецепторы дыхания аортальных и каротидных телец, чувствительные к гипоксии в артериальной крови, и барорецепторы стенки аорты и каротидных синусов, чувствительные к изменению системного артериального давления, расположены в рефлексогенных зонах в непосредственной близости друг от друга. Все названные рецепторы посылают афферентные сигналы к специализированным нейронам основного чувствительного ядра продолговатого мозга — ядра одиночного пучка. В непосредственной близости от этого ядра находится
дорсальное дыхательное ядро дыхательного центра. Здесь же в продолговатом мозге находится сосудодвигательный центр.
Координацию деятельности дыхательного и сосудодвигательного центров продолговатого мозга осуществляют нейроны ряда интег-ративных ядер бульбарной ретикулярной формации.
|
|