Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Взаимосвязь углеводного с другими видами обменов
|
|
Основная масса фосфорилированной глюкозы является для клеточных процессов энергоисточником (Рис. 3.34). Ее распад – гликолиз, протекающий в цитозоле, заканчивается (в аэробных условиях) пируватом, который, отправляясь в митохондрию, подвергается окислительному декарбоксилированию до ацетилКоА, часть же его молекул преобразуется в оксалоацетат. Затем оба вещества конденсируются в цитрат, который запускает цикл трикарбоновых кислот, поставляющий восстановительные эквиваленты – субстраты основных биоэнергетических процессов – биологического окисления и окислительного фосфорилирования.
Но для всех вышеописанных этапов разрушения глюкозо-6-фосфата существуют альтернативные ответвления. Метаболиты гликолиза (ГА-3-Ф, ДГАФ, ПВК) могут служить субстратами в синтезе некоторых заменимых аминокислот (ала, сер, гли, цис), а из дигироксиацетонфосфата путем его восстановления получается глицерол-1-фосфат – облигатный компонент глицерофосфатидов и особенно нейтральных жиров (в липоцитах). Продуктом трансаминирования ОА является важная для клеток аминокислота – аспартат.
При сдвиге баланса АДФ + Фн ↔ АТФ в правую сторону неиспользованный АТФ служит ингибитором ЦТК; накапливающийся цитрат выходит из матрикса митохондрии, распадается под действием АТФ-цитратлиазы на ОА и ацетилКоА. Последний путем конденсации способен превращаться или в холестерин, или в ацилы ВЖК, которые затем включаются в различные липиды: ТАГи, ГФ, СФ, ХС, ГЛ.
Возвращаясь к глюкозо-6-фосфату, следует заметить, что часть его пула может разрушаться по другому механизму – пентозофосфатному пути (ПФП). Его метаболиты – фосфорилированные пентозы, в первую очередь, рибозо-5-фосфат, которая затем включается в состав моно-, ди-, полинуклеотидов. Их представители выполняют самые различные функции, объединяя разные обмены. Среди простейших нуклеотидов выделяются макроэрги (АТФ, ГТФ, ЦТФ, УТФ), коферменты дегидрогеназ (ФМН), внутриклеточные гормоны (цАМФ, цГМФ). Динуклеотиды обычно входят в состав энзимов в виде коферментов (НАД+, НАД+Ф, ФАД – компоненты ДГ, модифицированный аналог НSКоА – участник трасацилирования). Нуклеиновые кислоты (РНК, ДНК) – полимеры, хранящие и передающие генетическую информацию. Другие фосфорилированные пентозы легко взаимодействуют с белками, липидами, меняя их свойства.
В процессе пентозофосфатного пути параллельно происходит восстановление НАД+Ф, но такая форма не идет (как НАДН) в митохондрии в ЭТЦ, а используется как источник водорода в реакциях синтеза ВЖК, холестерина, желчных кислот, стероидных гормонов, катехоламинов; в реакциях гидроксилирования аминокислот (фенилаланин → тирозин); в преобразовании рибонуклеотида в дезоксирибонуклеотид; является облигатным звеном в ферментативной антирадикальной защите; в гепатоцитах участвует в трансформации ксенобиотиков.
Но связи глюкозо-6-фосфата с другими звеньями метаболизма на этом не заканчиваются. Его небольшое количество (вклад зависит от вида ткани) взаимодействует с УТФ, образуя УДФ-глюкозу, которая или сама, или после модификации в другие моносахариды, включается в различные липиды (ГЛ) или биополимеры (полипептиды, белки). Окисление Г-6-A в УДФГК дает возможность получившемуся соединению также входить в состав вышеперечисленных структур, либо в гепатоцитах использоваться в процессах биотрансформации различных эндогенных веществ (билирубина) и ксенобиотиков.
Опасность возникновения гипогликемии (при голодании) в норме предотвращается за счет жизнедеятельности печени. В ее клетках хранятся гранулы гликогена, при необходимости распадающиеся до глюкозы. Но накопление данного гомополисахарида ограничено, поэтому, когда израсходуется это депо, вступает в действие более длительный источник эндогенной глюкозы – гликонеогенез. В нем могут участвовать представители самых различных классов, в том числе аминокислоты, глицерол, α -кетокислоты (Раздел III, Главы 2, 3), что служит еще одной иллюстрацией тесных связей между обменами (Рис. 3.32).
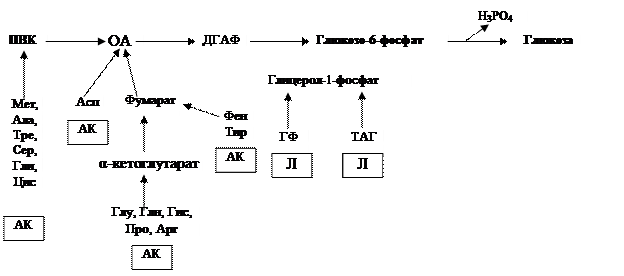
Рис. 3.32. Схема участия аминокислот и липидов в синтезе глюкозы в печени (ПВК – пируват, ОА – оксалоацетат, ДГАФ –дигидроксиацетонфосфат, АК – аминокислоты, ГФ – глицеролфосфат, ТАГ – триацилглицерол, Л - липиды)
|
|