Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Движение как средство взаимодействия организма с внешним миром. Строение мышечного волокна
|
|
Взаимодействие человека и животных с окружающим миром осуществляется через целенаправленную деятельность, или поведение. Важнейшим элементом поведения являются двигательные акты. За счет движения организма во внешней среде обеспечивается удовлетворение главных его потребностей. Ведущим фактором целенаправленного поведения, так же как и отдельного двигательного акта является получение полезного для жизнедеятельности организма приспособительного результата.
В процессе эволюции двигательные реакции существенно прогрессировали: от простого движения ложноножек у протистов до сложных двигательных актов млекопитающих и человека. При классификации движений учитывают основные функции, которые должна выполнять двигательная система. У человека таких функций в достаточно общей форме можно выделить несколько: 1) поддержание определенной позы и сохранение равновесия тела; 2) ориентация на источник внешнего сигнала для его наилучшего восприятия; 3) перемещение тела в пространстве (локомоция); 4) манипулирование; 5) мимика, жестикуляция; 6) речь, письмо.
Любой двигательный акт человека осуществляется благодаря сократительной деятельности скелетных мышц, управляемых центральной нервной системой. Структурной и функциональной единицей скелетной мышцы является поперечнополосатое мышечное волокно, имеющее диаметр от 10 до 100 мкм и длину 2–3 см. Каждое волокно – это многоядерное образование, возникающее в раннем онтогенезе, еще до рождения ребенка, из слияния клеток. Снаружи волокно покрыто оболочкой – сарколеммой. Внутри находится цитоплазма, называемая саркоплазмой. В саркоплазме расположены саркоплазматический ретикулум и сократительный аппарат мышечного волокна – миофибриллы. Миофибриллы имеют вид тонких нитей диаметром порядка 1 мкм, расположенных в саркоплазме вдоль волокна. В одном мышечном волокне может содержаться несколько сотен миофибрилл.
На всем своем протяжении миофибрилла состоит из повторяющихся участков – саркомеров длиной примерно по 2, 5 мкм. В световой микроскоп видно (рис. 6, А), что по краям саркомера расположены светлые участки, которые у соседних саркомеров смыкаются в I-диски, а в центре находится темный участок – А-диск. Саркомеры отделены друг от друга тонкими мембранами – Z-линиями, или Z-дисками. В поперечнополосатых (исчерченных) мышечных волокнах в соседних миофибриллах одноименные диски расположены на одном уровне, что придает волокнам регулярную поперечную полосатость (исчерченность).
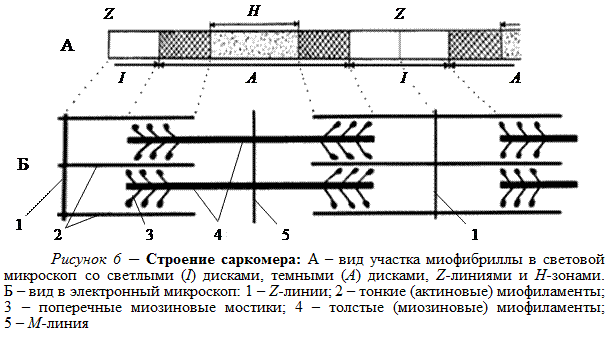
На электронных микрофотографиях видно (рис. 6, Б), что в темном А-диске расположены толстые, или миозиновые, нити (миофиламенты), состоящие из белка миозина и имеющие диаметр примерно 10 нм и длину 1, 6 мкм. Миозиновые миофиламенты имеют отходящие под углом выступы – поперечные мостики. На каждом миозиновом миофиламенте насчитывается около 500 мостиков. В середине саркомера видна тонкая темная М-линия, представляющая собой мембрану, которая, по-видимому, скрепляет вместе толстые нити.
В светлых I-дисках находятся тонкие миофиламенты, состоящие из белков актина, тропонина и тропомиозина. Актин составляет большую часть тонких миофиламентов и поэтому их называют актиновыми. Актиновые миофиламенты соседних саркомеров прикреплены к Z-линии.
Белки миозин и актин часто называют сократительными белками. Тропонин и тропомиозин в процессах сокращения и расслабления мышцы играют регуляторную роль.
 Миозиновые и актиновые миофиламенты расположены в саркомере параллельно, так, что тонкие нити могут скользить относительно толстых и заходить дальше в А-диск, что и происходит при сокращении мышцы. В расслабленной мышце концы тонких и толстых миофиламентов в малой степени перекрывают друг друга. Зона перекрывания в А-диске в световом микроскопе видна более темной (рис. 6 А), чем центральная Н-зона, в которой нет актиновых миофиламентов. При сокращении мышцы актиновые миофиламенты задвигаются между миозиновыми. Длина I-диска и Н-зона может уменьшаться до нуля, и саркомер становится короче.
Миозиновые и актиновые миофиламенты расположены в саркомере параллельно, так, что тонкие нити могут скользить относительно толстых и заходить дальше в А-диск, что и происходит при сокращении мышцы. В расслабленной мышце концы тонких и толстых миофиламентов в малой степени перекрывают друг друга. Зона перекрывания в А-диске в световом микроскопе видна более темной (рис. 6 А), чем центральная Н-зона, в которой нет актиновых миофиламентов. При сокращении мышцы актиновые миофиламенты задвигаются между миозиновыми. Длина I-диска и Н-зона может уменьшаться до нуля, и саркомер становится короче.
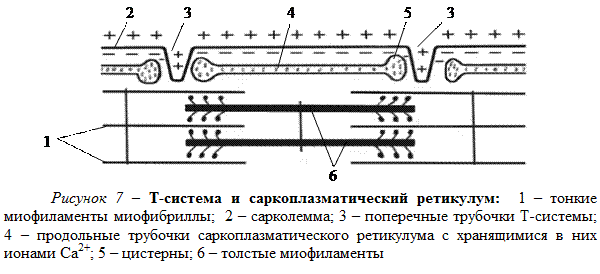 |
Сарколемма мышечного волокна имеет регулярные Т-образные впячивания (Т-система), имеющие вид поперечных трубочек, идущих вглубь волокна, перпендикулярно его продольной оси (рис. 7). Поперечные трубочки находятся в мышце млекопитающих примерно на границе А- и I-дисков и подходят к каждой миофибрилле.
Вдоль мышечного волокна в саркоплазме между миофибриллами расположены системы продольных трубочек саркоплазматического ретикулума. Каждая такая система продольных трубочек – это разветвленная, но замкнутая система, не соединяющаяся с внеклеточной средой. Продольные трубочки на концах имеют расширения в виде пузырьков (концевые цистерны). В трубочках с цистернами хранятся ионы Са2+. Концентрация Са2+ в трубочках и цистернах достигает 10-4 моль/л. Цистерны примыкают к поперечным трубочкам Т-системы.
Сократительные функции внутриклеточных органоидов, клеток, тканей и органов направлены на удовлетворение потребностей организма. Среди разнообразных форм сокращений ведущую роль играют мышечные сокращения. Главной особенностью скелетных мышц является то, что они в организме подчиняются произвольному контролю.
В настоящее время наиболее распространенной теорией, объясняющей процесс сокращения мышцы, является теория скольжения нитей (А. Хаксли и Д. Хансон). Согласно этой теории, сокращение мышцы обусловлено уменьшением длины саркомеров вследствие задвигания (скольжения) тонких миофиламентов дальше между миозиновыми. Механизм процессов, происходящих при мышечном сокращении следующий. Поступивший в нервно-мышечный синапс нервный импульс вызывает выделение из нервного окончания медиатора ацетилхолина. Ацетилхолин действует на сарколемму и изменяет ее проницаемость для ионов. В результате происходит изменение состояния сарколеммы и возникает специфический электрический заряд (потенциал действия), который распространяется по мембранам поперечных трубочек Т-системы вглубь мышечного волокна.
Предполагают, что поперечная трубочка образует электрический синапс на концевой цистерне продольной трубочки саркоплазматического ретикулума. Потенциал действия переходит на мембрану продольных трубочек, возбуждает ее, что в конечном итоге приводит к выходу ионов Са2+ из цистерн и трубочек в саркоплазму. В результате концентрация Са2+ в саркоплазме увеличивается с 10-8 моль/л до 10-6 моль/л. Ионы кальция проникают в миофибриллу и действуют на белок тропонин, входящий в состав тонких миофиламентов.
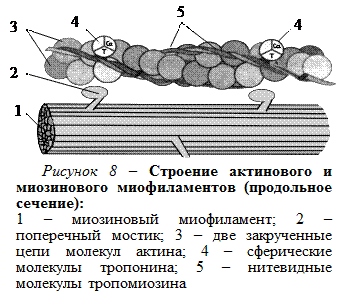 Тонкий миофиламент состоит из двух закрученных одна вокруг другой цепей шаровидных молекул актина (рис. 8). В желобках между цепями актина лежат нитевидные молекулы тропомиозина. Через одинаковые промежутки (примерно через 40 нм) на нитях тропомиозина располагаются сферические молекулы тропонина. Молекулы тропонина имеют тропомиозин-связывающие субъединицы, к которым прикреплены молекулы тропомиозина, и кальций-связывающие субъединицы.
Тонкий миофиламент состоит из двух закрученных одна вокруг другой цепей шаровидных молекул актина (рис. 8). В желобках между цепями актина лежат нитевидные молекулы тропомиозина. Через одинаковые промежутки (примерно через 40 нм) на нитях тропомиозина располагаются сферические молекулы тропонина. Молекулы тропонина имеют тропомиозин-связывающие субъединицы, к которым прикреплены молекулы тропомиозина, и кальций-связывающие субъединицы.
Если к мышце не поступают нервные импульсы и мышечное волокно не возбуждено, то миофибриллы находятся в расслабленном состоянии. Ионы кальция в это время находятся преимущественно в цистернах и трубочках саркоплазматического ретикулума, а в миофибриллах их концентрация очень мала. При практическом отсутствии ионов Са2+ в миофибрилле длинные молекулы тропомиозина располагаются так (рис. 9, А), что препятствуют контакту и прикреплению поперечных миозиновых мостиков к молекулам актина.
При возбуждении мышечного волокна вышедшие из продольных трубочек и проникшие внутрь миофибриллы ионы Ca2+ связываются с кальций-связывающей субъединицей молекулы тропонина. В результате молекула тропонина изменяет свое положение (поворачивается) и смещает в сторону нитевидные молекулы тропомиозина (рис. 9, Б). Устраняется препятствие для контакта поперечного миозинового мостика с молекулами актина. Миозиновый мостик соединяется сактином, образуя белок ак 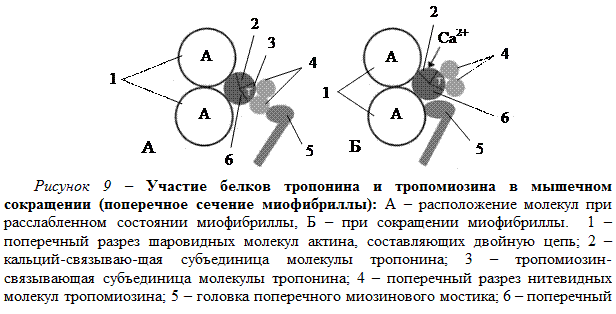
томиозин.
Образование актомиозина приводит к изменению конфигурации мостика – он сгибается и перемещает актиновый миофиламент (делает «гребок») на один шаг длиной примерно 20 нм (рис. 10).

В головках поперечных мостиков содержится фермент, который при контакте головки мостика с молекулой актина расщепляет АТФ, содержащейся в мышечном волокне. В результате высвобождается энергия. Высвободившаяся энергия израсходуется на «гребковые движения» мостиков и на работу «кальциевого насоса». «Кальциевый насос» начинает откачивать ионы Са2+ из миофибриллы обратно в цистерны саркоплазматического ретикулума.
Прямым источником энергии для мышечного сокращения является АТФ, без непосредственного соединения которой с миозиновыми мостиками не происходит «гребковых движений» и нет мышечного сокращения. Однако содержание АТФ в мышце невелико и достаточно для интенсивной работы примерно в течение 1 секунды. Предполагают, что на одно гребковое движение одного мостика тратится одна молекула АТФ.
Поэтому в мышце происходит постоянное восстановление или ресинтез АТФ, расщепившейся до АДФ. Энергия для ресинтеза АТФ выделяется в мышце в результате анаэробных и аэробных процессов. К анаэробным процессам относятся: 1) распад креатинфосфата (КФ); 2) бескислородное расщепление гликогена; 3) бескислородное расщепление глюкозы (гликолиз).
КФ– это первый энергетический резерв мышцы. При расщеплении КФ выделяется энергия, являющаяся быстрым источником восстановления АТФ. Запасы КФв мышце ограничены. Их достаточно для обеспечения работы мышцы примерно в течение 5 секунд. Энергия для ресинтеза КФи АТФ высвобождается при анаэробном расщеплении глюкозы и гликогена.
Бескислородное расщепление глюкозы и гликогена протекает в основном в тех случаях, когда сокращающиеся мышцы испытывают недостаток в снабжении кислородом. При этом образуется молочная кислота, что ведет к сдвигу рН в кислую сторону. По мере накопления молочной кислоты происходит угнетение активности (вплоть до полного торможения) ферментов и скорость гликолиза уменьшается. Поэтому при анаэробном расщеплении глюкозы и гликогена образуется не очень много АТФ.
Основное количество энергии, необходимой для ресинтеза АТФ, во время длительной работы при условии непрерывного поступления кислорода в митохондрии мышечных клеток высвобождается при аэробных процессах, т.е. при окислении белков, жиров и углеводов.
|
|