Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Биология Вып. 2
|
|
УДК 630.5(075), 630.653, 566+630*24
РАЗВИТИЕ ДРЕВОСТОЯ И ЕГО КОНСТАНТЫ
М. В. Рогозин, Г. С. Разин
Естественнонаучный институт Пермского государственного национального исследовательского университета, 614990, Пермь, ул. Генкеля, 4; zhal73@mail.ru; (342)2396722
В Пермском крае в моделях роста еловых древостоев обнаружены стабильные показатели – объем крон, наблюдаемый у древостоев с малой начальной густотой 1.0–1.3 тыс. шт./га, а в моделях любой начальной густоты – сбег ствола (отношение среднего диаметра к средней высоте древостоя за вычетом 1.3 м), которые неизменны только в пределах моделей с 45–50 и до 120 лет. Обсуждается концепция биополя и эпигенетические причины феномена биологической константы, а также диаметральных изменений производительности крон в редких и густых моделях на рубеже 40 лет. Подтверждена обоснованность гипотезы И.С. Марченко о существовании биополя древостоя, которое может определять развитие всего сообщества растений.
Ключевые слова: древостой; ход роста; модели развития; биопродуктивность; биополе.
О развитии древостоев с позиций разных наук
Развитие древостоев традиционно изучают лесоведение и лесная таксация, и в настоящее время обозначился кризис, сущность которого можно обозначить как отсутствие теории онтогенеза для сообщества древесных растений. Рассмотрим развитие дендроценоза пока в его наиболее простом виде, как развитие одноярусного древостоя от раннего возраста до возраста спелости.
 Кризис развивается давно. Еще в 1970-х гг. отмечалось, что в изучении причинных связей между древесным приростом и окружающей средой сделаны лишь первые шаги [Антанайтис, Загреев, 1981, с.70]. Тогда уже была известна теория динамики запаса древостоев Г.Ф. Хильми с формулой, имеющей три биофизические составляющие: поглощаемую листьями солнечную энергию, норму расхода солнечной энергии на поддержание жизнедеятельности и на прирост единицы запаса [Хильми, 1957].
Кризис развивается давно. Еще в 1970-х гг. отмечалось, что в изучении причинных связей между древесным приростом и окружающей средой сделаны лишь первые шаги [Антанайтис, Загреев, 1981, с.70]. Тогда уже была известна теория динамики запаса древостоев Г.Ф. Хильми с формулой, имеющей три биофизические составляющие: поглощаемую листьями солнечную энергию, норму расхода солнечной энергии на поддержание жизнедеятельности и на прирост единицы запаса [Хильми, 1957].
Ее теоретическая обоснованность несомненна, но сложность определения входящих в нее величин ограничивают и делают бесполезным ее применение в моделях хода роста [Верхунов, Черных, 2007]. Так, поглощаемая листьями солнечная энергия будет определяться всей фотосинтезирующей поверхностью листьев в пологе древостоя, которая теснее всего должна быть связана вначале с их массой и объемом и затем – с суммарным объемом крон, которые, в конечном счете, и производят древесину. Этот суммарный показатель еще только начинает изучаться.
Лесная таксация детально изучает лесную продукцию и традиционно ее интересует главным образом запас древесины, который появляется в результате работы полога древостоя. Но эта производящая древесину субстанция менее всего интересовала таксаторов и рассчитывалась иногда в виде объема сучьев и в редких случаях – в виде диаметров и объемов крон. По своему хозяйственному значению они стоят на последнем месте, но в биологическом смысле именно они – главные. Однако при изучении древесного прироста так вопрос даже не ставился; зависимость прироста древостоя от динамики объемов крон не изучалась, и тема прироста рассматривалась даже без увязки с ходом роста древостоев. Отмечалось, что такая увязка необходима при дальнейшем развитии учения о природе леса [Антанайтис, Загреев, 1981, с. 187].
Разработанные в эти годы модели прогноза прироста древостоя имели лаг прогноза в 5 лет и обеспечивали точность ± 5–8% при вероятности 68% [Никитин, Швиденко, 1978]. Они используются до сих пор и вполне приемлемы для учета лесов при корректировке их лесоустройством раз в 10 лет [Багинский, 2011]. Но если для отделов мониторинга и учета лесов такой короткий прогноз достаточен, то для лесовыращивания в конечном итоге нужен прогноз на 50–70 лет, при котором ошибки предложенных в 1970-е гг. моделей развития древостоев возрастут до 50–80%, и прогнозы, рассчитанные по таким моделям, становятся неприемлемыми. Дальнейшие исследования в лесной таксации были ориентированы в основном на развитие дистанционной инвентаризации лесов аэрокосмическими методами.
Сложность моделей с множеством переменных, действующих интегрально и меняющихся во времени, простирающемся за пределы жизни двух–трех поколений ученых, вынуждала изучать общее явление (развитие древостоя) по частям. При этом практика постоянно требовала применения полученных новых знаний. Общей теории онтогенеза никак не получалось [Шеверножук, 1980]. Но некоторые положения для такой теории в форме интегральных законов динамики древостоев нами разработаны [Разин, 1979; Разин, Рогозин, 2010].
Типы роста деревьев и древостоев, а также изменения за 40–60 лет бонитета насаждений на 2–3 класса нуждаются в отдельном обзоре; здесь лишь отметим, что селекционеры еще только начинают выяснять причины их появления в сомкнутых древостоях [Голиков, 2010], хотя предположения об их генетической обусловленности обсуждались уже давно [Кравченко, 1971; Кузьмичев, 1977; Котов и др., 1977]. Типы роста учитывают в сортоиспытаниях как неизбежность и поэтому отодвигают к возрасту спелости окончательные оценки элитности [Указания…, 2000]. Ученые-таксаторы первыми обнаружили типы роста и изучили их разнообразие, но до сих пор не могут объяснить причины их появления и поэтому не используют в моделировании роста древостоев, предпочитая более простые модели хода роста по классам бонитета [Багинский, 2011]. Неадекватность этих моделей мы отмечали неоднократно и объяснили появление типов роста различиями в начальной густоте, определяющей весь дальнейший ход роста древостоев [Разин, 1967; Разин, Рогозин, 2010; Рогозин, Разин, 2011].
В познании явления развития фитоценоза необходим принцип эмерджентности, то есть несводимости свойств целого к сумме свойств его частей. Соблюдение этого принципа невозможно в рамках какого-либо одного метода исследований. Поэтому отсутствие адекватных таблиц хода роста насаждений, как прогнозных моделей развития фитоценозов [Верхунов, Черных, 2007], вынуждает лесные науки все время искать эмпирические пути решений для регламента той или иной технологии выращивания леса. При этом в отсутствии общей теории развития древостоя, каждое направление ищет свои оптимальные параметры выращивания леса опытным путем, уходящим за пределы жизни ученого-исследователя. Так, появилось много работ, уточняющих и развивающих выдвинутые ранее идеи плантационного выращивания хвойных [Гаврилов, 1969; Шутов, 1985; Рябоконь, 1990]; однако по-прежнему нет работ, претендующих на обобщения, значимые для теории онтогенеза древостоев, которые могли бы быть использованы как при выращивании лесных культур, так и при рубках ухода. Частично они решены для молодняков с открытием «рангового закона роста деревьев в древостое» [Маслаков, 1980]. Однако неясно, как сохраняются ранги до возраста спелости, и поэтому остается пока в силе гипотеза о разных типах роста древостоев, хотя и без ответа на ключевой вопрос: какой процент молодых деревьев высших и низших рангов сохранили эти ранги до спелого возраста. Следует подчеркнуть, что без определения силы влияния конкуренции (густоты) на появление разных типов роста не может быть сделан и вывод об их генетической обусловленности, о которой иногда упоминают.
В работах по биопродуктивности лесов выяснены некоторые показатели полога древостоя, определяющие прирост леса. Так, в сосняках на Урале оказалась высокой связь диаметра дерева с массой хвои (ƞ =0.86–0.98) и общей массой кроны (ƞ =0.82–0.96). Запасы хвои после максимума снижаются и их динамика сходна с текущим приростом. Экстремум запаса хвои соответствует кульминации текущего прироста древостоя [Нагимов, Деменев, 1989].
Живой аппарат древостоя – фотосинтезирующая поверхность хвои, листьев и все клетки камбия – производит продукцию, и мы изучаем именно его результаты работы. Но сам этот аппарат оставлен лесоводами без внимания. Между тем вопросы, правильно заданные этому аппарату, могут дать ответы высокой эвристичности. Например, как оценивать и каким прибором «задать кроне дерева вопрос» о том, насколько она активна – будут весьма информативны, хотя окажутся и сложны в исполнении. Эти вопросы в целом связаны с концепцией биополя, введенной в биологию А.Г. Гурвичем [1945], который открыл митогенетическое излучение биообъектов. Источник поля А.Г. Гурвич связывал с центром клетки, позже с ядром, в конечном варианте теории – с хромосомами. Поле объекта в целом, согласно позднейшим представлениям, существует как сумма излучений полей всех его живых клеток. При этом поле имеет векторный, а не силовой характер [Гурвич, 1991].
Известная концепция конкуренции [Сукачев, 1953] не может объяснить все эффекты отношений внутри вида. В основе взаимоотношения растений со средой и между собой лежит материально-энергетический обмен, который чрезвычайно разнообразен – от обмена метаболитами и перераспределения элементов питания и воды до взаимодействия электрических полей, генерируемых растениями. Взаимодействие природных электромагнитных полей с полями растений имеет значение в информационных связях растений со средой и между растениями [Титов, 1978]. Эти поля дополняют концепцию фитогенного поля, предложенную А.А. Урановым [1965], где поля ограничивались только контурами растения – длиной корней, границами кроны. «В сообществе растений, как и во всякой другой системе осцилляторов, при достижении ими равновесного состояния неизбежно появление единой колебательной системы в результате настройки осцилляции отдельных растений на колебательный режим процессов обмена со средой, характерный для большинства взаимодействующих растений» [Титов, 1978, с. 126].
Убедительный и яркий пример такой настройки зафиксирован в 180-летних древостоях при изучения динамики образования годичного слоя древесины у ели и пихты на Урале. Оказалось, что деревья располагаются биогруппами и прирост рассредоточен во времени и по территории в трех ярусах полога; при этом деревья с близким типом прироста произрастают, как правило, на значительном расстоянии друг от друга. Отмечается, что «… внутренние причины, определяющие этот процесс, по силе влияния сопоставимы с внешними воздействиями» [Горячев, 1999]. Это явление можно интерпретировать как настройку деревьев друг относительно друга включением регуляторов роста, имеющих эпигенетическую природу и, возможно, в результате отбора в качестве соседей правых и левых форм по филлотаксису хвои. У этих форм различны предпочтения: левые формы предпочитают прямой свет и слабую конкуренцию, а правые – рассеянный и толерантны к конкуренции; установлено, что популяции представляют собой сложно структурированные системы с внутренней диссимметрической подразделенностью на левые и правые изопопуляции с зеркально-противоположными адаптивными свойствами [Голиков, 2010].
Используемые в физике методы обнаруживают сверхслабые излучения растений, но в полевых условиях они неприменимы. Но можно фиксировать реакцию растений на эти сигналы, в интерпретации И.С. Марченко [1995] «…по движению растений» (ветвей одного дерева относительно ветвей другого) и «растеканию растений в системе» (развитию полога древостоя); при этом малая величина излучения стимулирует заполнение экосистемы растениями, а большая – тормозит и «…предел растеканию наступает при максимуме биологического поля». Концепция биополя объясняет отворачивание побегов и растений друг от друга, колебания прироста в зависимости от сомкнутости, динамику изреживания, постоянство объема ветвей в единице объема полога древостоя и множество других явлений, не объяснимых с позиций известных физических и химических взаимодействий [Марченко, 1973, 1995].
Наличие экстремумов сомкнутости крон и пиков сомкнутости полога [Разин, 1979; Разин, Рогозин, 2009], а также совпадение максимумов текущего прироста и запасов хвои [Нагимов, Деменев, 1989] подтверждает основные законы экологии в заполнении пространства экосистемы растениями [Реймерс, 1994, с. 78]. Эти законы обусловлены действием физических, химических и энергетических взаимодействий растений (часть которых мы знаем как конкурентные). Можно предполагать, что механизмом саморегуляции в сообществе растений могут быть их биополя, излучающие сигналы самой различной физической природы, еще не изученные и во многом неизвестные.
Концепция биополя активно обсуждалась в 1970-е гг., и для древостоев были обнаружены доказательства его сильнейшего воздействия на прирост еловых культур еще до смыкания крон, хотя объяснение этому давалось другое [Кайрюкштис, Юодвалькис, 1976]: заблаговременное, за 2–3 года до смыкания крон, резкое снижение в 2 раза их прироста, начавшееся при расстоянии между кронами в 0.4 м; далее, после прорастания крон друг в друга на расстояние до 1 м, прирост плавно увеличивался в 1.6 раза относительно достигнутого минимума. Причины таких колебаний данные авторы объяснили «сменой внутривидовой конкуренции на взаимную толерантность», однако описать обнаруженное явление – еще не значит объяснить его причины. В настоящее время данное явление учитывают и для предотвращения торможения роста культур разреживания рекомендуются уже в самом раннем возрасте – в 9–13 лет [Большакова, 2007].
Поле биологического объекта по А.Г. Гурвичу существует как сумма полей его живых клеток. Биополе дерева в таком случае будут образовывать все его живые клетки, а именно клетки камбия и листвы (хвои), а сумма этих клеток у всех деревьев – биополе насаждения. Предполагается, что оно выступает как ведущий фактор естественного изреживания насаждений. Для биополя обнаружены и соответствующие константы, связанные с клетками камбия и листвы. Первая из них – объем ветвей в 1 м3 древесного полога (насыщенность сучьями). По мнению И.С. Марченко, она мало зависит от бонитета и в сомкнутых насаждениях есть величина постоянная; например, в зоне живой части крон в сосняках 1 бонитета эта величина от 30 до 90 лет изменялась в очень узких пределах от 793 до 815 см3/м3 [Марченко, 1995, с. 89].
Вторая константа – это предел насыщения полога клетками камбия в живой части кроны, измеряемая в см2/м3. Анализ молодняков сосны, осины, березы показал, что площадь клеток камбия стремится к пределу, после чего не увеличивается, и этот предел есть величина постоянная для каждой древесной породы. Поэтому «…каждая порода создает древесный полог с определенной напряженностью биологического поля» [Марченко, 1995, с. 86; с. 91].
Открытые И.С. Марченко константы не проверялись другими исследователями, и мы попытались выяснить их наличие в моделях роста еловых древостоев.
В качестве объектов исследования использованы 6 моделей роста естественных древостоев ели, составленных по вариантам начальной густоты в 10 лет в диапазоне от 7.9 до 1.0 тыс. шт./га для типов условий местопроизрастания С2–С3, помещенных в табличной форме в одной из наших работ [Разин, Рогозин, 2009]. Мы попытались выяснить наличие константных величин, анализируя развитие в пологе древостоя его основного показателя – суммы объемов крон. Для расчета использовали формулу
Vкр=Н× Скр× Lкр× 100k,
где Vкр – сумма объемов крон в пологе древостоя, м3/га;
Н – средняя высота, м;
Скр – сомкнутость крон, м2/м2;
Lкр – длина кроны в % от высоты ствола;
k – коэффициент, учитывающий форму кроны и плавно возрастающий от 0.332 в 10 лет до 0.354 в 120 лет.
Результаты и их обсуждение
Результаты расчетов суммы объемов крон (объем крон) в возрасте от 10 до 120 лет помещены в таблице и на рис. 1. Анализ данных показал, что в самых редких смолоду древостоях с начальной густотой 1.0 и 1.3 тыс. шт./га начиная с 40–45 лет объемы крон изменялись, соответственно, в пределах 50.6–52 и 46.4–47.5 тыс. м3/га с колебаниями от среднего уровня в пределах ± 1.4% (таблица). Колебания выглядят для этого возраста почти как прямая линия (рис. 1), их можно оценить как весьма незначительные, и начиная с 45 лет объем крон в таких древостоях можно считать величиной постоянной, то есть, в некотором смысле, «индивидуальной константой» для каждой модели древостоя.
При увеличении начальной густоты до 1.6 тыс. шт./га объем крон уже не так постоянен и колеблется в пределах 41–43.9 тыс. м3/га, при этом в 40 лет появляется слабый экстремум. При дальнейшем увеличении начальной густоты в моделях до 2.9–7.9 тыс. шт./га такой же слабый экстремум смещается к 30 годам, объемы неуклонно снижаются и достигают в 90–120 лет при начальной густоте 7.9 тыс. шт./га всего лишь 51% от объемов крон при начальной густоте 1.0 тыс. шт./га (таблица).
Сомкнутость крон (Скр) и сумма объемов крон (Vкр) в еловых древостоях в Пермском крае для типов условий местопроизрастания С2-С3
| Возраст, лет | Начальная густота в моделях роста деревостоев в 10 лет, тыс. шт./га | |||||||||||
| 1.0 | 1.3 | 1.65 | 2.9 | 5.1 | 7.9 | |||||||
| Скр, м² /м² | Vкр, м3/га | Скр, м² /м² | Vкр, м3/га | Скр, м² /м² | Vкр, м3/га | Скр, м² /м² | Vкр, м3/га | Скр, м² /м² | Vкр, м3/га | Скр, м² /м² | Vкр, м3/га | |
| 0.07 | 0.4 | 0.08 | 0.4 | 0.10 | 0.6 | 0.19 | 1.1 | 0.32 | 1.8 | 0.50 | 2.8 | |
| 0.20 | 2.3 | 0.24 | 2.8 | 0.31 | 3.6 | 0.54 | 6.2 | 0.89 | 10.1 | 1.18 | 12.1 | |
| 0.36 | 6.3 | 0.45 | 7.9 | 0.56 | 9.7 | 0.94 | 15.9 | 1.39 | 22.0 | 1.66 | 22.7 | |
| 0.58 | 13.8 | 0.70 | 16.5 | 0.87 | 20.2 | 1.30 | 28.0 | 1.59 | 29.3 | -1.61 | 24.8 | |
| 0.84 | 25.5 | 1.00 | 29.7 | 1.17 | 33.5 | 1.47 | 35.9 | 1.47 | 29.6 | -1.45 | 25.6 | |
| 1.08 | 38.6 | 1.21 | 41.4 | 1.33 | 42.5 | -1.38 | 35.8 | -1.33 | 29.7 | -1.3 | 25.5 | |
| 1.21 | 47.2 | 1.29 | 47.0 | 1.33 | 43.9 | -1.27 | 35.6 | -1.21 | 29.1 | -1.18 | 24.8 | |
| 1.25 | 50.6 | -1.28 | 47.5 | -1.27 | 43.5 | -1.20 | 35.3 | -1.13 | 28.4 | -1.09 | 24.0 | |
| -1.24 | 51.1 | -1.24 | 47.3 | -1.21 | 43.2 | -1.13 | 34.6 | -1.05 | 27.4 | -1.00 | 22.9 | |
| -1.17 | 50.7 | -1.15 | 47.1 | -1.11 | 42.6 | -1.02 | 33.2 | -0.93 | 25.6 | -0.87 | 20.9 | |
| -1.09 | 51.3 | -1.05 | 46.5 | -1.01 | 41.6 | -0.90 | 30.8 | -0.8 | 22.8 | -0.73 | 18.0 | |
| -1.05 | 51.7 | -1.01 | 46.4 | -0.97 | 41.3 | -0.85 | 29.9 | -0.75 | 21.8 | -0.67 | 16.8 | |
| -1.03 | 52.0 | -1.00 | 47.1 | -0.95 | 41.2 | -0.84 | 29.9 | -0.73 | 21.5 | -0.65 | 16.5 | |
| Примечание. Знак «минус» в значении сомкнутости крон означает, что пик сомкнутости пройден и значения снижаются. |
Как видим, сумма объемов крон елового фитоценоза оказывается величиной постоянной только в пределах некоторых линий развития, определяемых начальной густотой и только для самых редких смолоду древостоев (рис. 1). Эти постоянные величины имеют ясный биологический смысл, как предел, больше которого полог уже не может заполняться биомассой и сохраняет ее на фоне изменения буквально всех таксационных показателей, включая даже класс бонитета, который у моделей с начальной густотой 1 – 1.3 тыс. шт./га с 40 до 120 лет меняется на 0.6 класса – от 1.2 до 1.8 [Разин, Рогозин, 2009].
В то же время если рассчитать производительность работы крон, измеряя ее текущим общим приростом древесины по десятилетиям, то на 1000 м3 объема крон густые модели 5 и 6 производят в 20-30 лет на 7 – 21% больше древесины в сравнении с моделями 1 – 3 с малой густотой. Но в 40 лет во всех моделях она выравнивается, а далее меняется на обратную: в 60 – 110 лет кроны в более редких моделях 1 – 3 снижают производительность в 1.4 – 1.8 раза в сравнении с моделью 6; в 120 лет эти различия почти исчезают (рис. 2). То есть в густых моделях до 40 лет производительность работы крон оказывается ниже, а после 40 лет – выше, чем в редких моделях. Данное явление тем более удивительно и необычно на фоне неуклонного снижения конечных показателей продуктивности и запаса в густых моделях.
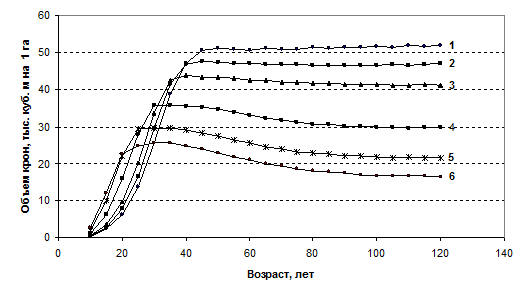
Рис. 1. Динамика объема крон в еловых древостоях с начальной густотой в 10 лет:
1 – 1.0; 2 – 1.3; 3 – 1.6; 4 – 2.9; 5 – 5.1; 6 – 7.9 тыс. шт./га
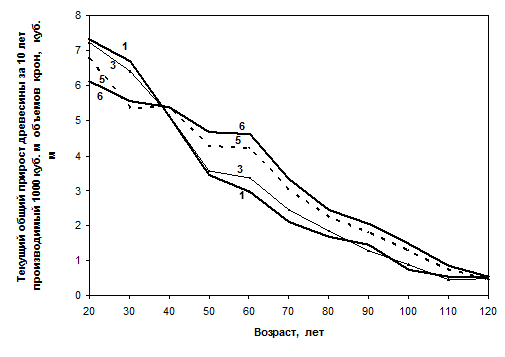
Рис. 2. Динамика производительности крон в моделях еловых древостоев с начальной густотой:
1 – 1.0; 3 – 1.65; 5 – 5.1; 6 – 7.9 тыс. шт./га
Производительность работы крон в динамике в разных по густоте древостоях как явление совершенно не изучалось ранее. Следует отметить, что выравнивания данных, полученных для настоящей статьи, мы не проводили, и на рис. 2 видны случайные пересечения линий в 90–110 лет у соседних моделей 1 и 3, а также в 30 лет – у моделей 5 и 6, что было бы обязательно устранено аппроксимацией, если бы при составлении моделей роста [Разин, Рогозин, 2009] этот показатель учитывали.
Мы можем только предполагать почему наблюдаются такие разные линии динамики производительности крон. Их можно объяснить, например, отбором в густых ценозах правых форм растений, более продуктивных при повышении плотности популяции [Голиков, 2010], включением эпигенетических регуляторов роста и формированием биогрупп с разной динамикой прироста [Горячев, 1999], а также грузом старой теневой хвои, которая сохраняется у ели до 7–9 лет. В редких древостоях ее больше, тогда как в густых старая хвоя отмирает быстрее, и поэтому эффект работы фотосинтезирующего аппарата повышается. Возможна и гипотеза И.С. Марченко, совершенно по-новому объясняющая это явление, согласно которой крона имеет 4 слоя по активности клеток камбия и листвы; активность этих слоев определяет взаимодействие биополя дерева с соседями, а прирост ветвей зависит не только от нахождения кроны в тени или на свободе, но и от малоизученных пока явлений усиления и торможения роста клеток в этих слоях кроны [Марченко, 1995].
Какой из приведенных четырех весьма сложных процессов превалирует и в каком возрасте – пока совершенно неясно даже предположительно.
Среди других признаков, претендующих на название «биологическая константа», мы обнаружили показатель, известный как сбег ствола дерева (отношение диаметра на высоте 1.3 м к высоте дерева). В моделях поведение этого признака вполне предсказуемо и в возрасте после 30–50 лет может быть отнесено к стабильному: для 7 выбранных для сравнения моделей его величина устойчива на протяжении 70–90 лет, плавно повышаясь к концу периода всего на 2–4%, тогда как различия между моделями за это время составляют 5–7% (рис. 3). Для представления о поведении этого признака в особенно густых древостоях на рисунке помещена дополнительно модель с густотой 14 тыс. шт./га.
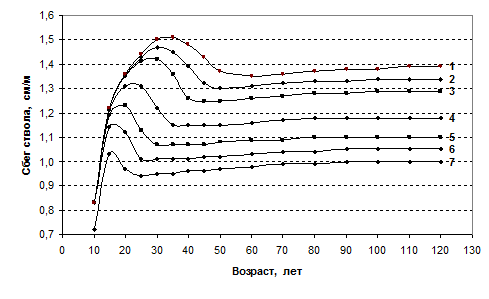
Рис. 3. Динамика сбега ствола в моделях еловых древостоев с начальной густотой:
1 – 1.0; 2 – 1.3; 3 – 1.65; 4 – 2.9; 5 – 5.1; 6 – 7.9; 7 – 14 тыс. шт./га
Еще лучший результат получается, если в показатель внести поправку и рассчитать сбег по отношению к высоте ствола выше измерения на нем диаметра, т.е. отношением диаметра к высоте с вычетом высоты груди (минус 1.3 м). Полученные линии динамики несколько иные, но поведение сбега ствола в них начиная с 45 – 50 лет выглядит как несомненно стабильное (рис. 4).
Следует отметить, что в специальных работах по изучению формы стволов [Кофман, 1986] подобных закономерностей не обнаружено, так как изучались не естественные ряды древостоев, а их предельные состояния в разном возрасте и в древостоях, разных по начальной густоте.
Сбег ствола имеет глубокий биологический смысл для дерева и характеризует последствия его развития в условиях конкуренции. При сильном давлении (большой густоте) сбег мал, а в условиях свободы развития он увеличен и деревья более «коренастые». Его определяет степень развития кроны – чем она больше, тем больше и сбег, и если мы говорим о типах, линиях и моделях роста и их изменении регулированием плотности стояния деревьев рубками, то быстрый отклик, и изменение размеров кроны и сбега ствола возможны до начала его стабилизации; если же сбег стабилизировался, то преодолеть его инерцию, имеющую физиологическую и эпигенетическую природу, будет, скорее всего, невозможно, так как программа развития дендроценоза уже наполовину реализована и кроны развитых деревьев сформировали устойчивую форму.
Кульминацию развития насаждения в лесоводстве и таксации обычно связывают с максимумом текущего прироста запаса. В наших моделях текущие приросты по запасу выглядят как серия пересекающихся кривых и каждая из них имеет свой максимум (рис. 5). Максимумы наблюдаются в редких моделях в 30–40, а в густых – в 25 лет, и они случились за 15 лет до возраста стабилизации сбега ствола. Если же древостой перешел свой предел по приросту, то есть миновал его кульминацию, обусловленную множеством причин, вызвавших именно такое развитие древостоя, то «переместить» развитие древостоя на более производительную по этому показателю линию разреживанием трудно или почти невозможно в силу отмеченной выше инерции развития.
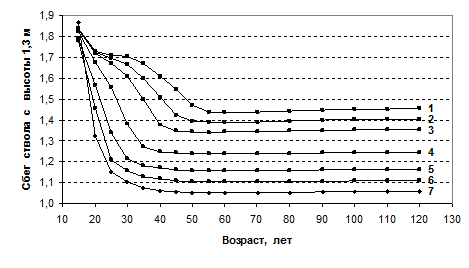
Рис. 4. Динамика сбега ствола с высоты 1.3 м в моделях еловых древостоев с начальной густотой:
1 – 1.0; 2 – 1.3; 3 – 1.65; 4 – 2.9; 5 – 5.1; 6 – 7.9; 7 – 14 тыс. шт./га
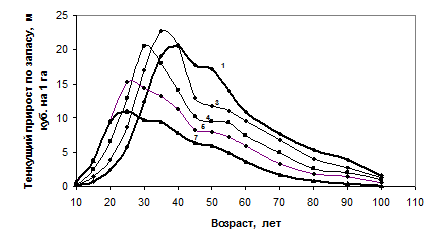
Рис. 5. Текущий прирост по запасу в моделях еловых древостоев с начальной густотой:
1 – 1.0; 3 – 1.65; 4 – 2.9; 5 – 5.1; 7 – 14 тыс. шт./га
Отсюда следует и важнейшее практическое правило: регулирования текущей густоты (рубки ухода «прочистки» и «прореживания») должны быть завершены примерно за 15–20 лет до начала стабилизации сбега ствола и в период до кульминации текущего прироста по запасу. Это будет возраст 25 лет в густых моделях развития (начальная густота 5.1 тыс. шт./га и более) и 30–35 лет – в редких моделях роста (начальная густота 2.9 тыс. шт./га и менее).
Возможно, именно по причине запоздалых прочисток и прореживаний в опытах С.Н. Сеннова было получено парадоксальное заключение о том, что рубки ухода не повышают общую производительность древостоев [Сеннов, 1984], что совершенно не согласуется с современными результатами выращивания леса после ранних изреживаний [Рябоконь, 1990; Большакова, 2007].
Полученные нами результаты рождают скорее больше вопросов, чем ответов. Важно далее эти вопросы «правильно задать» древостою. По-видимому, биоконстанту для дендроценозов следует искать в показателях, характеризующих полог древостоя. Необходимо изучение и такого трудного явления, как биополе дендроценоза. В соответствии с концепцией биополя, возможно, будут найдены некоторые стабильные биологические и биофизические признаки и, скорее всего, они будут подчинены энергетической константе этого поля.
Затронутые в настоящее статье вопросы особенно актуальны для лесной селекции, так как невозможность прогноза роста деревьев и древостоев сдерживает использование полученных в 1 – 2 классах возраста оценок роста потомства популяций и плюсовых деревьев. Нахождение постоянных величин в развитии древостоев, своеобразных биологических и биофизических констант, поможет раскрыть механизм появления восходящих, нисходящих и стабильных типов роста деревьев и древостоев, найти периоды максимума показателей, необходимых для моделирования и прогноза роста древостоев.
Следует отметить, что мы изучили динамику показателя суммы объемов живых крон, а он отличается от изученных И.С. Марченко двух других показателей: объема ветвей в 1 м3 полога (насыщенности сучьями) и площади клеток камбия в живой части кроны. Эти показатели мы не могли проверить, но гипотеза И.С. Марченко о существовании этих показателей в виде постоянных величин находит свое косвенное подтверждение в развитии крон у древостоев ели в самых редких по начальной густоте вариантах моделей роста.
Полог древостоя, несомненно, имеет предел в заполнении пространства листвой, и этот предел определяет динамику и функциональную организацию всего сообщества и отдельных растений в нем. Именно изучение динамики крон и полога позволило разработать методику моделирования [Разин, 1979] и модели хода роста, которые инициировали открытие первых 5 законов в теории естественной динамики древостоев [Разин, Рогозин, 2010, 2012].
Заключение
В Пермском крае в моделях роста еловых древостоев для типов условий местопроизрастания С2 – С3 обнаружены стабильные показатели – объем крон, наблюдаемый у древостоев с малой начальной густотой 1.0 – 1.3 тыс. шт./га, а в моделях любой начальной густоты – сбег ствола (отношение среднего диаметра к средней высоте древостоя за вычетом 1.3 м), которые неизменны только в пределах моделей начиная с 45 – 50 и до 120 лет. Объемы крон составляют 50.6 – 52 м3/га при начальной густоте 1.0 тыс. шт./га и 46.4 – 47.5 м3/га при начальной густоте 1.3 тыс. шт./га. Производительность работы крон в моделях с большей начальной густотой до 40 лет оказывается ниже, а далее – выше, чем в редких моделях. Для объяснения феномена биологической константы и диаметральных изменений производительности крон в редких и густых моделях на рубеже 40 лет важно вскрыть физиологические, эпигенетические и биополевые причины этих явлений.
Для нахождения «главной константы» необходимо изучение биополя дендроценоза. Вероятно, она будет представлять собой предельное значение интенсивности излучений конгломерата биологических полей всех деревьев в единице объема полога, и именно она будет определять все развитие древостоя. Интенсивность биополя будет зависеть, возможно, от преобладания правых или левых форм растений, имеющих различия в характере фотосинтеза. Можно полагать, что в соответствии с концепцией биополя будут найдены стабильные признаки, подобные обнаруженному нами объему крон, и они будут подчинены некоторой и пока еще гипотетической энергетической константе.
Библиографический список
Антанайтис В.В., Загреев В.В. Прирост леса. М.: Лесн. пром-сть, 1981. 200 с.
Багинский В.Ф. Ход роста древостоев и его отражение в таблицах и математических моделях // Лесн. хоз-во. 2011. № 2. С. 40 – 42.
Большакова Н.В. Влияние густоты и размещения посадочных мест на рост ели при выращивании культур по интенсивным технологиям: автореф. дис. … канд. с-х. наук. СПб., 2007. 24 с.
Верхунов П.М., Черных В.Л. Таксация леса. Йошкар-Ола: МарГТУ, 2007. 395 с.
Гаврилов Б.И. Лесные плантации быстрого роста // Лесн. журн. 1969. № 4. С. 14 – 16.
Голиков А.М. Теоретическое и прикладное значение эколого-дисимметрийного подхода в исследовании формовой и генетической структуры популяций видов хвойных // Наука о лесе ХХI в.: материалы междунар. конф. Гомель, 2010. C. 157 – 160.
Горячев В.М. Влияние пространственного размещения деревьев в сообществе на формирование годичного слоя древесины хвойных в южнотаежных лесах Урала // Экология. 1999. № 1. С. 9 – 19.
Гурвич А.Г., Гурвич Л.Д. Митогенетическое излучение. М.: Медгиз, 1945.
Гурвич А.Г. Принципы аналитической биологии и теории клеточных полей. М.: Наука, 1991.
Кайрюкштис Л.В., Юодвалькис А.И. Явление смены характера взаимоотношений между индивидами внутри вида // Лесоведение и лесное хозяйство. Минск, 1976. Вып. II. С. 16 – 24.
Котов М.М. и др. Типы деревьев сосны по росту в высоту и их значение в семеноводстве // Лесн. журн. 1977. № 6. С. 27 – 30.
Кофман Г.Б. Рост и форма деревьев. Новосибирск: Наука, 1986. 210 с.
Кравченко Г.Л. Этапы онтогенеза сосны обыкновенной // Лесоведение. 1971. № 6. С. 17 – 20.
Кузьмичев В.В. Закономерности роста древостоев. Новосибирск: Наука, 1977. 158 с.
Маслаков Е. Л. Об особенности роста и дифференциации деревьев в молодняках сосны // Восстановление и мелиорация лесов Северо-Запада РСФСР. Л.: ЛенНИИЛХ, 1980. С. 53 – 61.
Никитин К.Е., Швиденко А.З. Методы и техника обработки лесоводственной информации. М., 1978. 164 с.
Марченко И.С. Биополе лесных экосистем. Брянск: БТИ, 1973. 91 с.
Марченко И.С. Биополе лесных экосистем. Брянск: БГИТА, 1995. 188 с.
Нагимов З.Я., Деменев В.В. Особенности формирования надземной фитомасы сосновых древостоев на Среднем Урале // Проблемы использования, воспроизводства и охраны лесных ресурсов: материалы респ. науч.-практ. конф. Йошкар-Ола, 1989. С. 28 – 29.
Разин Г.С. О бонитетных шкалах и ходе роста древостоев // Сб. тр. / Поволжский ЛТИ им. М. Горького. Йошкар-Ола, 1967. № 58. Вып. 3. С. 101 – 105.
Разин Г.С. Динамика сомкнутости одноярусных древостоев // Лесоведение. 1979. № 1. С. 23 – 25.
Разин Г.С., Рогозин М.В. О ходе роста древостоев. Догматизм в лесной таксации // Вестн. Перм. ун-та. 2009. Вып. 10(36). Биология. С. 9 – 38.
Разин Г.С., Рогозин М.В. О законах и закономерностях роста и развития, жизни и отмирания древостоев // Лесн. хоз-во. 2010. № 2. С. 19 – 20.
Разин Г.С., Рогозин М.В. Теория естественной возрастной динамики одноярусных древостоев // Лесн. хоз-во. 2012. № 3. С. 41 – 42.
Рогозин М.В., Разин Г.С. Лесные культуры Теплоуховых в имении Строгановых на Урале: история, законы развития, селекция ели. Пермь, 2011. 192 с.
Реймерс Н.Ф. Экология (теории, законы, правила, принципы и гипотезы). М.: Журнал «Россия молодая», 1994. 367 с.
Рябоконь А.П. Продуктивность сосновых насаждений и качество древесины в них при ускоренном выращивании на пиловочник и балансы // Лесн. журн. 1990. № 6. С. 19 – 24.
Сеннов С.Н. Уход за лесом (экологические основы). М.: Лесн. пром-сть, 1984. 128 с.
Сукачев В.Н. О внутривидовых и межвидовых взаимоотношениях среди растений // Сообщ. ин-та леса. 1953. Вып. 1. С. 5 – 44.
Титов Ю.В. Эффект группы у растений. Л.: Наука, 1978. 151 с.
Указания по лесному семеноводству в Российской Федерации / Федер. служба лесн. хоз-ва России. М., 2000. 197 с.
Уранов А.А. Фитогенное поле // Проблемы соврем. ботаники. М.; Л.: Наука, 1965. Т. 1. С. 251 – 254.
Хильми Г.Ф. Теоретическая биогеофизика леса. М., 1957. 205 с.
Шеверножук Р.Г. Ранняя диагностика в лесной селекции. Воронеж, 1980. 52 с. Деп. в ЦБНТИлесхоз 8.12.1980, № 60 лд.
Шутов И.В. Лесосырьевые плантации ели и сосны // Лесн. хоз-во. 1985. № 3. С. 34 – 37.
Поступила в редакцию 07.05.2012
Development of the forest stand and its constant
M. V. Rogozin, candidate of agricultural sciences, associate professor
G. S. Razin, consultant
Natural science institute of Perm state university. 4, Genkel str., Perm, Russia, 614990; zhal73@mail.ru; (342)2396722
In Perm Krai in models of growth of fir-tree forest stands stable indicators – the volume of kroner observed at forest stands with small initial density of 1.0–1.3 thousand pieces/hectares, and in models of any initial density – taper a trunk (the relation of average diameter to average height of a forest stand minus 1.3 m) which are invariable only within models from 45–50 and till 120 years are found. The concept of a biofield and the epigenetichesky reasons of a phenomenon of a biological constant, and also diametrical changes of productivity of kroner in rare and dense models at a boundary in 40 years is discussed. Validity of a hypothesis of I.S. Marchenko about existence of a biofield of a forest stand which can define development of all community of plants is confirmed.
Key words: forest stand; course of growth; development model; bioproductivity; biofield.
Рогозин Михаил Владимирович, кандидат сельскохозяйственных наук, зав. лабораторией
Разин Геннадий Сергеевич, консультант
Естественнонаучный институт ФГБОУВПО «Пермский государственный национальный исследовательский университет»
|
|