| 1.Энергетический метаболизм. Сущность.
Способы получения энергии у микроорганизмов различны. Некоторые микроорганизмы (фототрофы) способны перерабатывать химическую энергию солнечного света, другие (хемотрофы) получают энергию путем окисления химических веществ.
Окислением принято считать процесс отнятия двух атомов водорода. Этот процесс носит название дегидрирование. Восстановление того или иного соединения представляет собой присоединение двух атомов водорода (гидрирование).
Окисление может быть представлено следующим образом:
АН2 ® А + 2Н+,
В + 2Н+ ® ВН2 + А
Суммарное уравнение: АН2 + В ® BH2 + А
В этой реакции АН2 – восстановитель или донор ионов водорода, а В – окислитель (акцептор), так как присоединяет ионы водорода.
Водород и электроны, отнятые от окисляемого субстрата (донора), переносятся к конечному акцептору не непосредственно, а ступенчато, поэтапно с помощью окислительно-восстановительных ферментов. К таким ферментам относятся дегидрогеназы, которые переносят водород, и ферменты цитохромной системы – цитохромы и цитохромоксидаза, которые переносят электроны. Дегидрогеназы и цитохромная система образуют дыхательную цепь.
Набором окислительно-восстановительных ферментов объясняется отношение микроорганизмов к молекулярному кислороду. В зависимости от способа получения энергии и от конечного акцептора водорода микроорганизмы можно разделить на три физиологические группы:
• облигатные аэробы – микроорганизмы, которые не могут существовать без кислорода. Энергию эти микроорганизмы получают в результате окисления веществ в присутствии кислорода воздуха (дыхания). У этих микроорганизмов в клетках имеется полный набор окислительно-восстановительных ферментов, осуществляющих перенос протона водорода и электронов на кислород. Примером микроорганизмов этой группы являются микроскопические грибы и уксуснокислые бактерии;
• облигатные анаэробы – микроорганизмы, для которых кислород является клеточным ядом. Такие микроорганизмы получают энергию в процессе брожения. В составе клеток облигатных анаэробов имеются специфические дегтдрогеназы и отсутствуют цитохромы и цитохромоксидаза. Представителями облигатных анаэробов являются маслянокислые бактерии рода Clostridium, бифидобактерии;
• факультативные анаэробы – микроорганизмы, способные жить как в присутствии кислорода, так и без него (осуществляют как дыхание, так и брожение). В клетках факультативных анаэробов присутствуют кроме окислительно-восстановительных ферментов, переносящих водород и электроны на кислород (участвуют в процессе дыхания) специфические ферменты (участвуют в процессе брожения). К факультативным анаэробам относятся дрожжи, молочнокислые, пропионовокислые бактерии и другие микроорганизмы.
Энергия, образуемая при энергетическом обмене, трансформируется в энергию макроэргических связей молекул АТФ. Процесс образования АТФ называется фосфорилированием.
Механизм образования АТФ у разных групп микроорганизмов неодинаков. Поэтому различают субстратное, окислительное и фотофосфорилирование.
Фотофосфорилирование – образование АТФ при поглощении квантов света молекулами хлорофилла. В результате от молекулы хлорофилла отрываются электроны, которые, проходя по цепи переноса электронов, отдают свою энергию системе АДФ-АТФ, в результате чего энергия света трансформируется в энергию макроэргических связей АТФ.
Субстратное фосфорилирование – образование АТФ непосредственно на молекуле субстрата. Протекает в анаэробных условиях на стадиях превращения 1, 3-дифосфоглицериновой кислоты в 3-фосфоглицериновую кислоту и фосфоэнолпировиноградной кислоты в пировиноградную кислоту (в процессе гликолиза).
Окислительное фосфорилирование – образование АТФ одновременно с процессом переноса протонов и электронов по дыхательной цепи ферментов. На каждые 2 атома водорода, поступившие в дыхательную цепь, синтезируются 3 молекулы АТФ. Окислительное фосфорилирование осуществляется аэробными и факультативно-анаэробными микроорганизмами.
2.Цитоплазма клетки, пластиды.
Пластиды. В цитоплазме клеток всех растений находятся пластиды. В клетках животных пластиды отсутствуют. Различают три основных типа пластид: зеленые - хлоропласты; красные, оранжевые и желтые - хромопласты; бесцветные - лейкопласты.
Хлоропласт. Эти органоиды содержатся в клетках листьев и других зеленых органов растений, а также у разнообразных водорослей. Размеры хлоропластов 4-6 мкм, наиболее часто они имеют овальную форму. У высших растений в одной клетке обычно бывает несколько десятков хлоропластов. Зеленый цвет хлоропластов зависит от содержания в них пигмента хлорофилла. Xлоропласт - основной органоид клеток растений, в котором происходит фотосинтез, т. е. образование органических веществ (углеводов) из неорганических (СО2 и Н2О) при использовании энергии солнечного света.
По строению хлоропласты сходны с митохондриями. От цитоплазмы хлоропласт отграничен двумя мембранами - наружной и внутренней. Наружная мембрана гладкая, без складок и выростов, а внутренняя образует много складчатых выростов, направленных внутрь хлоропласта. Поэтому внутри хлоропласта сосредоточено большое количество мембран, образующих особые структуры - граны. Они сложены наподобие стопки монет.
В мембранах гран располагаются молекулы хлорофилла, потому именно здесь происходит фотосинтез. В хлоропластах синтезируется и АТФ. Между внутренними мембранами хлоропласта содержатся ДНК, РНК. и рибосомы. Следовательно, в хлоропластах, так же как и в митохондриях, происходит синтез белка, необходимого для деятельности этих органоидов. Хлоропласты размножаются делением.
Хромопласты находятся в цитоплазме клеток разных частей растений: в цветках, плодах, стеблях, листьях. Присутствием хромопластов объясняется желтая, оранжевая и красная окраска венчиков цветков, плодов, осенних листьев.
Лейкопласты. находятся в цитоплазме клеток неокрашенных частей растений, например в стеблях, корнях, клубнях. Форма лейкопластов разнообразна.
Хлоропласты, хромопласты и лейкопласты способны клетка взаимному переходу. Так при созревании плодов или изменении окраски листьев осенью хлоропласты превращаются в хромопласты, а лейкопласты могут превращаться в хлоропласты, например, при позеленении клубней картофеля.
Яндекс.ДиректВсе объявленияИщите в сети с Яндекс.Браузером! Поиск по интернету из адресной строки! Нужные сайты всегда под рукой.browser.yandex.ru 0+
Аппарат Гольджи. Во многих клетках животных, например в нервных, он имеет форму сложной сети, расположенной вокруг ядра. В клетках растений и простейших аппарат Гольджи представлен отдельными тельцами серповидной или палочковидной формы. Строение этого органоида сходно в клетках растительных и животных организмов, несмотря на разнообразие его формы.
В состав аппарата Гольджи входят: полости, ограниченные мембранами и расположенные группами (по 5-10); крупные и мелкие пузырьки, расположенные на концах полостей. Все эти элементы составляют единый комплекс.
Аппарат Гольджи выполняет много важных функций. По каналам эндоплазматической сети к нему транспортируются продукты синтетической деятельности клетки - белки, углеводы и жиры. Все эти вещества сначала накапливаются, а затем в виде крупных и мелких пузырьков поступают в цитоплазму и либо используются в самой клетке в процессе ее жизнедеятельности, либо выводятся из нее и используются в организме. Например, в клетках поджелудочной железы млекопитающих синтезируются пищеварительные ферменты, которые накапливаются в полостях органоида. Затем образуются пузырьки, наполненные ферментами. Они выводятся из клеток в проток поджелудочной железы, откуда перетекают в полость кишечника. Еще одна важная функция этого органоида заключается в том, что на его мембранах происходит синтез жиров и углеводов (полисахаридов), которые используются в клетке и которые входят в состав мембран. Благодаря деятельности аппарата Гольджи происходят обновление и рост плазматической мембраны.
Лизосомы. Представляют собой небольшие округлые тельца. От Цитоплазмы каждая лизосома отграничена мембраной. Внутри лизосомы находятся ферменты, расщепляющие белки, жиры, углеводы, нуклеиновые кислоты.
К пищевой частице, поступившей в цитоплазму, подходят лизосомы, сливаются с ней, и образуется одна пищеварительная вакуоль, внутри которой находится пищевая частица, окруженная ферментами лизосом. Вещества, образовавшиеся в результате переваривания пищевой частицы, поступают в цитоплазму и используются клеткой.
Обладая способностью к активному перевариванию пищевых веществ, лизосомы участвуют в удалении отмирающих в процессе жизнедеятельности частей клеток, целых клеток и органов. Образование новых лизосом происходит в клетке постоянно. Ферменты, содержащиеся в лизосомах, как и всякие другие белки синтезируются на рибосомах цитоплазмы. Затем эти ферменты поступают по каналам эндоплазматической сети к аппарату Гольджи, в полостях которого формируются лизосомы. В таком виде лизосомы поступают в цитоплазму.
Клеточный центр. В клетках животных вблизи ядра находится органоид, который называют клеточным центром. Основную часть клеточного центра составляют два маленьких тельца - центриоли, расположенные в небольшом участке уплотненной цитоплазмы. Каждая центриоль имеет форму цилиндра длиной до 1 мкм. Центриоли играют важную роль при делении клетки; они участвуют в образовании веретена деления.
Клеточные включения. К клеточным включениям относятся углеводы, жиры и белки. Все эти вещества накапливаются в цитоплазме клетки в виде капель и зерен различной величины и формы. Они периодически синтезируются в клетке и используются в процессе обмена веществ.
Ядро. Каждая клетка одноклеточных и многоклеточных животных, а также растений содержит ядро. Форма и размеры ядра зависят от формы и размера клеток. В большинстве клеток имеется одно ядро, и такие клетки называют одноядерными. Существуют также клетки с двумя, тремя, с несколькими десятками и даже сотнями ядер. Это - многоядерные клетки.
Ядерный сок - полужидкое вещество, которое находится под ядерной оболочкой и представляет внутреннюю среду ядра.
Преподаватель: _________ О.В. Кузнецова
|


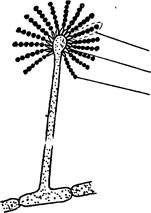 а) б)
Рис. 4.2 - Строение плодоносящих гифов:
а – спорангиеносца, б – конидиеносца
2.Классификация вирусов, строение, формы.
формы, химический состав вирусов и фагов. Классификация вирусов
Вирусная частица (вирион) состоит из спирально закрученной нуклеиновой кислоты – ДНК или РНК, покрытой снаружи белковой оболочкой (капсидом). Капсид состоит из отдельных субъединиц – капсомеров, которые идентичны друг другу.
Содержание нуклеиновой кислоты и белка у разных вирусов неодинаковое. Так, у вируса гриппа на долю нуклеиновой кислоты приходится 1 % (по массе), у вируса полиомиелита –25%, у бактериофагов – 50–60% белка.
При исследовании вирусов под электронным микроскопом обнаружены следующие формы вирусов:
• палочковидная (вид прямого цилиндра). Такую форму имеет вирус табачной мозаики;
• нитевидная (изгибающиеся эластичные нити). Эту форму имеют вирусы некоторых растений;
• сферическая. Такую форму имеет вирус гриппа, герпеса;
• октаэндрическая (форма многогранника). Это вирус полиомиелита, вирус полиомы, аденовирусы;
• булавовидная (головастикообразная, сперматозоидная). Такую форму имеют вирусы бактерий – бактериофаги (рис. 5.1).
а) б)
Рис. 4.2 - Строение плодоносящих гифов:
а – спорангиеносца, б – конидиеносца
2.Классификация вирусов, строение, формы.
формы, химический состав вирусов и фагов. Классификация вирусов
Вирусная частица (вирион) состоит из спирально закрученной нуклеиновой кислоты – ДНК или РНК, покрытой снаружи белковой оболочкой (капсидом). Капсид состоит из отдельных субъединиц – капсомеров, которые идентичны друг другу.
Содержание нуклеиновой кислоты и белка у разных вирусов неодинаковое. Так, у вируса гриппа на долю нуклеиновой кислоты приходится 1 % (по массе), у вируса полиомиелита –25%, у бактериофагов – 50–60% белка.
При исследовании вирусов под электронным микроскопом обнаружены следующие формы вирусов:
• палочковидная (вид прямого цилиндра). Такую форму имеет вирус табачной мозаики;
• нитевидная (изгибающиеся эластичные нити). Эту форму имеют вирусы некоторых растений;
• сферическая. Такую форму имеет вирус гриппа, герпеса;
• октаэндрическая (форма многогранника). Это вирус полиомиелита, вирус полиомы, аденовирусы;
• булавовидная (головастикообразная, сперматозоидная). Такую форму имеют вирусы бактерий – бактериофаги (рис. 5.1).
