Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Дыхательная цепь. Биоэнергетика.
|
|
Основной источник энергии для всех организмов на Земле — солнечное излучение (в результате реакций ядерного синтеза на Солнце). Под действием солнечной энергии путем фотосинтеза в клетках растений электроны в молекулах Н2О возбуждаются, т. е. переходят на более высокий энергетический уровень и включаются в состав белков, липидов, углеводов. В процессе распада белков, жиров и углеводов в организме животных совершается обратный переход электронов на более низкий энергетический уровень с образованием Н2О, что сопровождается высвобождением такого же количества энергии. Следовательно, основной носитель энергии — электрон, а ее источник – Солнце.
Существуют различные формы энергии в природе (тепловая, химическая, электрическая, механическая и др.), взаимосвязь которых отражается законами термодинамики.
Условия, необходимые для сохранения гомеостаза живых организмов (постоянства внутренней среды) – это поступление в неё энергии, т.к. в тканях непрерывно совершаются процессы распада веществ, высвобождения и расхода энергии. В организм животных энергия поступает в форме белков, углеводов и липидов, катаболизм которых ведет к освобождению из них энергии и трансформации ее в энергию макроэргов (АТФ и др.), электрическую энергию, тепловую энергию, механическую, энергию химических связей и др.
Реакции рефосфорилирования АДФ и последующего использования АТФ в качестве источника энергии (дефосфорилирования) образуют цикл, который повторяется 2, 5-3 тыс. раз в сутки. Существует 2 способа рефосфорилирования АДФ или синтеза АТФ: окислительное фосфорилирование, субстратное фосфорилирование. Основные субстраты для рефосфорилирования АДФ: окисление белков, жиров и углеводов в ходе их катаболизма в тканях.
Оксидазный путь окисления белков, липидов, углеводов (тканевое дыхание) – основной путь окисления в тканях у животных (используется около 90% кислорода, поступающего в клетки) и одновременно производства энергии (АТФ и тепла). Оксидазный путь окисления субстратов обеспечивается ферментами и коферментами дыхательной цепи. Из них формируется цепь переноса электронов, и кислород используется ими только в качестве акцептора электронов, а затем протонов. Компоненты дыхательной цепи (все они сложные белки, за исключением убихинона) являются энзимами и полипептидами, которые содержат в качестве простетической группы ряд коферментов и кофакторов, из которых формируются редокс-пары, расположенные в определенном порядке (в порядке возрастания их редокс-потенциалов). Редокс-потенциал (окислительно-восстановительный потенциал) характеризует способность переносчика принимать или отдавать электроны.
Компоненты дыхательной цепи в митохондриях организованы в комплексы: четыре белковых комплекса (I, III, IV, V), встроенных во внутреннюю митохондриальную мембрану и две подвижные системы (молекулы-переносчики): – убихинон (КоQ) и цитохромы с. Сукцинатдегидрогеназа из ЦТК рассматривается как комплекс II дыхательной цепи. Электроны и протоны, снимаемые с белков, липидов, углеводов (реакции дегидрирования), поступают в дыхательную цепь двумя путями:
1) при дегидрировании субстратов с помощью НАД-зависимых дегидрогеназ комплекс I переносит электроны и протоны через ФМН и FeS-белок на убихинон;
2) при окислении субстратов ( сукцината) флавинзависимыми дегидрогеназами электроны и протоны переносятся на убихинон комплексом II, содержащим ФАДН2-дегидрогеназу и FeS-белок. Далее электроны попадают на убихинон и систему цитохромов и только цитохром а3 (в составе цитохромоксидазы) может передать электроны на 1/2О2 с образованием ионов О-2, последние соединяются с протонами, снятыми с окисляемых субстратов через дегидрогеназы и KoQ. Образуется эндогенная или метаболическая вода (за сутки образуется 300 – 400 мл). Перенос электронов по дыхательной цепи сопровождается постепенным освобождением из них энергии, часть которой (~ 40%) используется на образование АТФ, а остальная энергия рассеивается в виде тепла (теплопродукция). Энергия электронов используется, прежде всего, для переноса протонов в межмембранное пространство митохондрий и формирования протонного градиента (электрохимического потенциала) на внутренней митохондриальной мембране. Образование АТФ сопряжено с обратным потоком протонов из межмембранного пространства в матрикс митохондрии. Однако мембрана митохондрий непроницаема для протонов. В митохондриях только АТФ-синтетаза (комплекс V) позволяет осуществить обратное движение протонов из межмембранного пространства в матрикс митохондрий, и этот же энзим катализирует образование АТФ. Таким образом, синтез АТФ сопряжен с окислением субстратов, а затем коферментов и кофакторов дыхательной цепи с участием кислорода. Поэтому процесс (окисления субстратов и фосфорилирования АДФ с образованием АТФ) получил название окислительного фосфорилирования. Сопряжение окисления и фосфорилирования (синтез АТФ) возможны только на 3 или 2 участках дыхательной цепи, где энергии выделяется в достаточном количестве для фосфорилирования АДФ и образования АТФ.
Если протоны поступают в межмембранное пространство через комплексы I, III и IV, то образуется 3 молекулы АТФ и коэффициент фосфорилирования (отношение 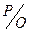 ) = 3; если протоны поступают через комплексы II, III и IV, то образуется 2 молекулы АТФ и коэффициент фосфорилирования () = 2. Коэффициент фосфорилирования () – это количество неорганического фосфата, включенного в 2 или 3 молекулы АТФ в процессе переноса 2 e- на 1 атом кислорода. При некоторых условиях окисление, т.е. перенос электронов по дыхательной цепи, происходит, но без синтеза АТФ (фосфорилирования). Этот процесс называется разобщением окислительного фосфорилирования. Разобщители окислительного фосфорилирования (протонофоры, термогенины, свободные жирные кислоты и др.) способствуют расходованию электрохимического потенциала в обход АТФ-синтетазы и поэтому АТФ не образуется. В этих условиях дыхание усиливается, фосфорилирование подавляется (снижается образование АТФ), но увеличивается теплопродукция. Скорость образования АТФ зависит от энергетического состояния клеток, т.е отношения: [АТФ] / [АДФ] * [Фн]. При использовании организмом энергии часть АТФ гидролизуется до АДФ и Фн, энергетический заряд клетки снижается. Повышение концентрации АДФ автоматически увеличивает скорость окислительного фосфорилирования и образования АТФ, т.е. с помощью АДФ контролируется дыхание митохондрий. Этот механизм регуляции энергетического обмена клетки получил название дыхательный контроль. Увеличение содержания АДФ приводит к ускорению и дыхания, и фосфорилирования.
) = 3; если протоны поступают через комплексы II, III и IV, то образуется 2 молекулы АТФ и коэффициент фосфорилирования () = 2. Коэффициент фосфорилирования () – это количество неорганического фосфата, включенного в 2 или 3 молекулы АТФ в процессе переноса 2 e- на 1 атом кислорода. При некоторых условиях окисление, т.е. перенос электронов по дыхательной цепи, происходит, но без синтеза АТФ (фосфорилирования). Этот процесс называется разобщением окислительного фосфорилирования. Разобщители окислительного фосфорилирования (протонофоры, термогенины, свободные жирные кислоты и др.) способствуют расходованию электрохимического потенциала в обход АТФ-синтетазы и поэтому АТФ не образуется. В этих условиях дыхание усиливается, фосфорилирование подавляется (снижается образование АТФ), но увеличивается теплопродукция. Скорость образования АТФ зависит от энергетического состояния клеток, т.е отношения: [АТФ] / [АДФ] * [Фн]. При использовании организмом энергии часть АТФ гидролизуется до АДФ и Фн, энергетический заряд клетки снижается. Повышение концентрации АДФ автоматически увеличивает скорость окислительного фосфорилирования и образования АТФ, т.е. с помощью АДФ контролируется дыхание митохондрий. Этот механизм регуляции энергетического обмена клетки получил название дыхательный контроль. Увеличение содержания АДФ приводит к ускорению и дыхания, и фосфорилирования.
Кроме оксидазного, существуют другие варианты окисления веществ в тканях. Оксигеназный способ окисления субстратов катализируется энзимами моно- и диоксигеназами, которые включают один или 2 атома кислорода, соответственно, в субстрат. Таким способом окисляются желчные кислоты, стероидные гормоны, обезвреживаются чужеродные вещества в организме. В процессе перекисного окисления липидов (ПОЛ) реакционно-способные свободные радикалы окисляют ненасыщенные жирные кислоты, что приводит к образованию гидропероксидов липидов, спиртов, альдегидов, кетонов, малонового диальдегида. ПОЛ стимулирует обновление биологических мембран, выполняет бактерицидную функцию, но также активирует разрушение белков, углеводов, липидов, мембран. Антиоксидантные системы организма защищают клетки от ПОЛ. При пероксидазном окислении веществ образуется перекись водорода, которая обезвреживается ферментами каталазой или пероксидазой.
|
|