Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Биологические функции полисахаридов
|
|
Поскольку полисахариды в тканях и биологических жидкостях находятся только в связанном состоянии с белком, то правильнее рассматривать биологическое значение преимущественно углеводного компонента в тех углевод-белковых комплексах, в которых на его долю приходится большая часть. Такие углевод-белковые комплексы называют протеогликанами в отличие от гликопротеидов, у которых углеводный фрагмент меньше, чем белковый, или имеет некоторые особенности (см. гл. «Смешанные макромолекулы»). К наиболее важным функциям полисахаридов отйосятсй: энергетическая, опорная, защитно-механическая, связующая (структурная), гидроосмотическая и ионрегулирующая, кофакторная.
Энергетическую функцию выполняют резервные полисахариды — крахмал и гликоген. Они являются депо углеводов в клетке и при необходимости быстро расщепляются на легко усвояемый источник энергии — глюкозу. Гликоген и крахмал пищи выполняют ту же функцию после обработки их ферментами пищеварительного тракта.
Опорную функцию выполняют структурный полисахарид целлюлоза в растительных организмах и хондроитинсульфаты в костной ткани.
Защитно-механическая — типичная функция кислых гетерополисахарн- дов. Высокая вязкость и слизеподобная консистенция кислых гетероиоли- сахаридов объясняет их роль биологического смазочного материала, защищающего поверхность клеток. Выстилая трущиеся поверхности сосудов, мочеполовых путей, пищеварительного тракта, слизистой носа, трахеи, бронхов, суставов (синовиальная жидкость) и т. д.,. они предохраняют их от механического повреждения. Механическую устойчивость внутренним органам при сжатии и вибрации придают межклеточные-гелеподобные мукополиса- хариды; у водных растений они обусловливают упругость и прочность при ударах воды.
Связующая, или структурная, функция — кислые гетерополисахариды являются структурным межклеточным веществом, одновременно выполняющим функцию биологического цемента (например, гиалуроновая кислота).
Гидроосмотйческая и ион регулирующая функции. Кислые гетерополисахариды благодаря высокой гидрофильности и отрицательному заряду способны удерживать большие количества воды н катионов. Например, гиалуроновая кислота — чрезвычайно гидрофильный полисахарид. Она связывает межклеточную воду и катионы, регулируя межклеточное осмотическое давление. Подобно осмометру эта кислота препятствует излишнему скоплению свободной воды в межклеточном пространстве. Избыточное связывание некоторых ионов может привести к нарушениям, например, к кальцификации сосудистой стенки.
Кофакторная функция. Некоторые гетерополисахариды, такие как гепарин и гепарансульфат, действуют как кофакторы ферментов. При соединении с белком они образуют активный белок — полисахаридный комплекс. Гепарин проявляет свойства тех ферментных белков, у которых он играет роль кофактора. Поэтому у него проявляется антисвертывающая функция (задерживает свертывание крови) и Энтилипемическая (снижает уровень липидов в крови, активируя их расщепление). Антикоагулирующими свойствами обладает и дерматансульфат, который содержится в аорте.
На практике гепарин и сульфатированные синтетические полисахариды (гепариноиды) широко применяют как антикоагулянты и противоатероскле- ротическне (препятствующие отложению лнпидов в сосудах) препараты.
ГЛАВА 5. НУКЛЕИНОВЫЕ КИСЛОТЫ
Нуклеиновые кислоты были впервые выделены швейцарским ученым Ф. Ми- " шером (1868) из ядер клеток гноя. Он показал, что это соединение содержит фосфор, и назвал его нуклеином (от лат. nucleus.— ядро). По современным понятиям это был нуклеопротеид. В 1872 г. Мишер использовал другой материал, богатый ядрами, — сперму лососей — для выделения нуклеиновых кислот. И вновь им была получена нуклеиновая кислота в соединении с веществом основного характера, получившим название протамин. Впоследствии из объектов животного, растительного мира и микроорганизмов были выделены разные нуклеиновые кислоты. Наилучшим источником нуклеиновых кислот оказались клетки, имеющие большое ядро, поэтому широко использовалась вилочковая железа (тимус). Гидролиз нуклеиновой кирлоты из тимуса показал, что в ней содержатся пуриновые и пнримидиновые основания (аденин, гуанин, цитозин, тимин), 2-дезокси-0-рибоза и фосфорная кислота. Эту кислоту назвали дезоксирибонуклеиновой (ДНК)- Из дрожжей была получена другая нуклеиновая кислота, содержащая вместо тимина урацил и вместо дезоксирибозы рибозу. Ее назвали рибонукленновЬй кислотой (РНК)- Лишь в начале 40-х годов нашего столетия было доказано, что РНК тоже является обязательным компонентом всех животных, растительных и бактериальных клеток.
1. Общая характеристика нуклеиновых.кислот
Нуклеиновыми кислотами или полинуклеотидами называются высокомолекулярные вещества, ' состоящие из мононуклеотидов, соединенных в цепь З'.б-фосфодиэф ирными связями.
Общее содержание ДНК и РНК в клетках зависит от их функционального состояния. В сперматозоидах количество ДНК достигает 60% (в пересчете на сухую массу клеток), в большинстве клеток 1 — 10, а в мышцах около 0, 2%. Содержание РНК, как правило, в 5—10 раз больше, чем ДНК- Соотношение РНК/ДНК в печени, поджелудочной железе, эмбриональных тканях и вообще в тканях, активно синтезирующих йелок, составляет от 4 до 10. В тканях с умеренным синтезом белка соотношение колеблется от 0, 3 до 2, 5. Особое место занимают вирусы. У них в качестве генетического.материала может быть либо ДНК (ДНК-овые вирусы), либо РНК (РНК-овые вирусы).
В клетках бактерий, не имеющих ядра (прокариоты), молекула ДНК (хромосома) находится в специальной зоне цитоплазмы — нуклеоиде. Если она связана с клеточной мембраной бактерии, то ее называют мезосо- мой. Фрагмент ДНК меньших размеров локализуется вне этой хромосомной зоны. Такие участки ДНК в бактериях называются, плазмидами или эписомами. В клетках, имеющих ядро (эукариоты), ДНК распределена между ядром, где она входит в состав хромосом и ядрышка, и внеидерными органоидами (митохондриями и хлоропласта ми). Имеются наблюдения, что в очень малых количествах ДНК присутствует в микросомах. Однако это еще требует доказательств.
Примерно 1 — 3% ДНК клетки приходится на внеядерную ДНК, а остальное сосредоточено в ядре. Значит, наследственные свойства характерны не только для ядра, но и для митохондрий и хлоролластов клеток. Необычно высоким содержанием внеядерной ДНК отличаются зрелые яйцеклетки, у которых она присутствует в многочисленных митохондриях и желточных пластинках, причем в последних является не генетическим материалом, а резервом нуклеотидов.
РНК в отличйе.от ДНК распределена по клетке более равномерно. Уже одно это обстоятельство говорит о том, что функция РНК более динамична и многообразна. В клетках высших организмов около 11% всей РНК находится в ядре, около 15% —в митохондриях, 50% —в рибосомах и 24% — в гиалоплазме. 70
Молекулярная масса ДНК зависит от степени сложности живого объекта: у бактерий она составляет 2 • 10s, у человека и животных достигает 10". У бактерий ДНК находится в виде единичной гигантской молекулы, слабо связанной с белками. В других объектах ДНК окружена белками или простейшими аминами. У вирусов это простейшие основные белки или полиамины (путресцин я спермидин), которые нейтрализуют отрицательный заряд молекулы ДНК, связываясь с ее фосфатными группами., В - сперматозоидах некоторых животных и рыб ДНК образует комплексы с протаминами и гистоноподобнымн белками. В хромосомах клеток человека и других высших организмов ДНК связана с гистонами и негистоновыми белками. Такие комплексы белка с ДНК называют дезоксирибонуклеопротеидами (ДНП).
РНК имеет значительно меньшую молекулярную массу, чем ДНК- В зависимости от выполняемой функции, молекулярной массы и состава нуклеотидов выделяют следующие главные типы РНК: информационная, или матричная (мРНК), транспортная (тРНК) и рибосомальная (рРНК). Разные рРНК различаются по молекулярной массе (табл. 13). Кроме трех
Таблица 13. Краткая характеристика нуклеиновых кислот клеток высших организмов
|
главных типов есть минорные, или редкие, РНК, содержание которых в клетке незначительно, и функции их только изучаются.
Большинство типов РНК связано в клетке с различными белками. Такие комплексы называются рибонуклеопротеидами (РНП). Характеристика нуклеиновых кислот суммирована в табл. 13.
2. Компоненты нуклеиновых кислот Выделенные и очищенные ДНК и РНК гидролизуют в специальном режиме кислотами (ДНК) или щелочами (РНК); чаще гидролиз проводят специаль-
Нуклеопротенды i
{-------
дкп
(Дезок сирибонуклеопротенд)
протамнны и яр.)
| ---- 1 РНП {Рибону югеопроте ид) h' |
ДНК
Главные мононуклеотиды - (дезоксирибоиуклеотнды)
д-АМФ д-ГМФ д-ЦМф д-ТМФ
Главные мононуклеотиды (рибонуклеотнды)
АМФ ГМФ ЦМФ УМФ
X
Дезок сир ибоиук леознцы д-Аденозмн д-Гуаиозин д-Цитндии ц-Тимишн
XI
Г
| ~1 Н, Р04 |
| f н, го4 ...ГЦ | Рибоаа |
N-Основаиия Аденин Гуанин Цитоэии
Рнбонуклеоэиды Аде но энн Гуанозия Цитидин Урнднн —t—
Ы-0 снования Аденци Гуанин Цитоэии; Урация;
Рис. 7. Схема разделения нуклеиновых к
ными ферментами — нуклеазами, которые разрывают фосфодиэфирные связи в полинуклеотидах. При гидролизе фосфоднэфнрных связей нуклеиновых кислот освобождаются их структурные мономеры — мононуклеотиды, которые состоят из азотистого основания, пентозы и фосфорной кислоты {рис. 7).
Азотистые основания
Азотистые основания нуклеиновых кислот делятся по химическому строению на две группы — пуриновые и пиримидиновые. Среди них имеются главные и редкие (минорные) пуриновые и пиримидиновые основания. К главным пуриновым относятся аденин и гуанин; к пиримидиновым — цитозин, урацил и тимин. В ДНК входят аденин, гуанин, цитознн и тимнн, а в РНК — все те же основания, за исключением тимина, который замёнен урацилом. Главные основания имеют следующее строение:
Nf^CH
14 5 14' Ln П1» un " И V™'V.
«У»
(5-м егклурядил)
I ' I
*fh< ^x N ^VX HM -V\
ll л»сн СН I У с
Важной особенностью их является способность к лактам-лактимной -таутомерии. Например:
но о........
ноАЛ
У
Минорные основания встречаются главным образом в тРНК и в следовых количествах в рРНК. К ним относятся дополнительно метилированные пуриновые и пиримидиновые основания (например, 2-метиладенин, {-метилгуанин, 5-метилцитозин, 5-оксиметилцитозин н др.).
Нуклеозиды
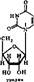
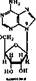
Вещества, в которых азотистые основания соединен^ с пентозой, называются нуклеозидами. Нуклеозиды относятся к N-гликоЗкдам'. У них атом, С-1 пентозы связан с N-3 пурина или N-1 пиримидина. Ъ зависимости от типа пентозы различают два вида нуклеозидов — дезоксирибонуклеозиды, содержащие 2-дезоксирибозу, и рибонуклеозиды, содержащие рибозу:
NH,
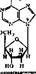
|
|
|
Дезоксирибонуклеозиды входят только в ДНК, а рибонуклеозиды — только в РНК. Пиримидиновые и пуриновые-нуклеозиды содержат соответствующие азотистые основания:,
Рибонуклеозиды
Аденнк + рибоза — яденозии Гуакнн + рнбоэа — гуанознн ЦНтозхн + рнбоэа — цетклин Ураикл +■ рнбоэа — урнднн
Д еэоксирибоиукдеои иды
Аденнн + дезоксирибоза — дезоксиадепоэин Гуаннн+дезоксиркбоза — дезокснгуанознн Цитознн + дезоксирибоза — дезоксииитидин Тниин + дезоксирибоза — дезокситииндин
Кроме' главных встречаются минорные нуклеозиды, в которые входят минорные азотистые основания. Больше всего минорных нуклеозидов содержится в тРНК. Наиболее распространенными минорными нуклеозидами, входящими во все тРНК, являются дигидроуридин, псевдоуридин {обозначаемый сокращенно значком и риботимидин. В псевдоуридине отсутствует обычная N-гликозидная связь. В нем атом С-1 рибозы соединен с атомом С-5 урацила.
Нуклеотиды
Нуклеотиды представляют собой соединения соответствующего типа нуклеози- да с фосфорной кислотой. Они также делятся на рибонуклеотиды, содержащие рибозу, и дезоксирибонуклеотиды, содержащие 2'-дезоксирибозу. Ниже приведено строение адениловых нуклеотидов.
| . нА н |
| НО—Р—О—СН, |
| НО—Р—О—СН- 8 ■ Кн н- |
Фосфат может присоединяться в разные положения кольца пентоза {в рибонуклеотидах — в положениях 2', 3', 5', в дезоксирнбонуклеотидах — в положения 3', 5'). Имеющиеся в клетке свободные нуклеотиды содержат фосфатную группу в" положении 5'. Нуклеознд-5'-фосфаты участвуют в биологическом синтезе; нуклеиновых кислот и образуются при их распаде. Поскольку нуклеозид-5'-фосфаты, или моно'нуклеотиды, являются производными соответствующих куклеозидов, то различают те же главные и редкие рибомоно- нА
| O-iUJ- |
снош н- 5' -моЪюфоефцт
нуклеотнды и дезоксирибомононуклеотнды. Удлинение фосфатного конца мо- нонуклеотида за счет присоединения дополнительных фосфатов приводит к образованию нуклеозидполифосфатов:
—I—О—СН- Ан '
т-5'-чонофоеф»т (АМФ) i-js-iwфосфат (ад»)
5ИН-5'-трифосф1Г (АТФ)
Чаще Всего в клетках встречаются нуклеозиддифосфаты и нуклеозидтрифос- фаты. Ниже приводятся названия и сокращенные обозначения нуклеознд- фос фатов:
Рибонуклеоэидфосфаты ю-, дн-, трифосфат «о-, ли-, трифосфат ю-, ди-, трифосфат «о-, ди-, трифосфат
| Аденоэин Гуанозии Цитидня Урнднн |
| АМ4>, АДФ, АТФ ГМФ, ГДФ, ГТФ ЦМФ, ЦДФ. ЦТФ УМФ, УДФ. УТФ |
| Дезокснаденозин Д езокси'гу а нознн Дезоксицитидии Дезокситнмидин |
| дАМФ, дАДФ, дАТФ дГМФ, дГДФ, дГТФ дЦМФ, дДДФ. дЦТФ дТМФ, дТДФ, дТТФ |
Дезоксирибомухлеоэ идфосфаты MOHO-, ди-, трифосфат НОИО-, дн-, трифосфат иоио-, ди-, трифосфат моно-. ди-. трифосфат
Все нуклеоз ид фосфаты находятся в клетке в виде анионов, поэтому аденсх- зинфосфаты правильнее обозначать АМФ2-, АДФ3-, АТФ4~. АДФ и АТФ являются макроэргическими, т. е. богатыми энергией, соединениями, химическая энергия которых используется организмом для различных функций. Остальные нуклеозидди- и трифосфаты также участвуют в реакциях синтеза биологических веществ.
Производные нуклеозидов. Среди производных нуклеотндов можно выделить циклические нуклеотиды {например, 3', 5'-АМФ и 3', 5'-ГМФ), которые являются универсальными регуляторами обмена вещества внутри клетки. Большая группа производных нуклеотндов служит коферментами различных Ферментов, участвуя в реакциях превращения веществ. Имеются кофермен- ты — производные моионуклеотидов и динуклеотидов (см. гл. 10 «Ферменты»).
3. Строение н уровни организации нуклеиновых кислот
Нуклеиновые кислоты имеют первичную, вторичную и третичную структуру.
|
|