Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Слухова сенсорна система
|
|

5.3. Дослідження основних зорових функцій.
Дослідження розпочинають з визначення гостроти зору, або бінокулярного зору. При аналізі результатів дослідження звертають увагу на фокусування зображення на сітківку ока та механізми формування рецепторного потенціалу у паличках або колбочках.
У паличках зоровий пігмент родопсин розташований у їх зовнішньому сегменті у мембрані дисків, у його складі є білок скотопсин і альдегід вітаміну А – ретиналь. Коли світло падає на сітківку ретиналь родопсину переходить із форми 11 цис-ретиналь у повний транс-ретиналь, який через серію проміжних речовин перетворюється на метародопсин ІІ, який активує на мембрані дисків G-білок, що має назву трансдуцин, який в свою чергу активую фермент фосфодіестеразу. Фосфодіестераза каталізує перетворення цГМФ до 5-ГМФ, а концентрація цГМФ зменшується, що призводить до закривання воріт натрієвих кналів, наслідком чого є виникнення гіперполяризації у фоторецепторах.

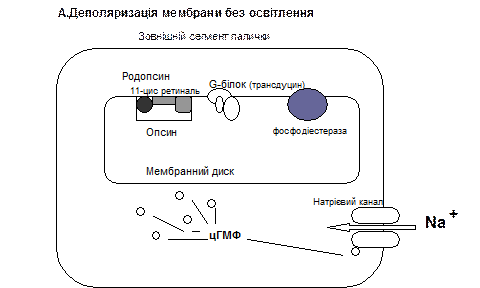

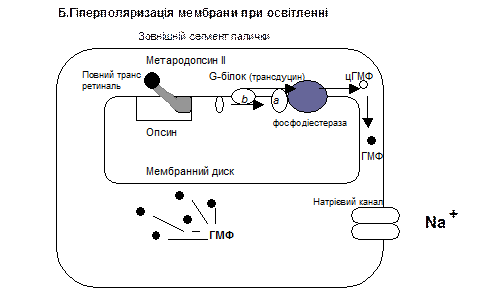
Гіперполяризація мембрани фоторецептора призводить до зменшення виділення гальмівного медіатора, завдяки чому виникає деполяризація біполярних клітин сітківки, які передають інформацію на гангліонарні клітини, аксони яких утворюють зоровий нерв, в якому генеруються ПД.
Світло у фоторецепторах викликає каскад реакцій, що знижують концентрацію внутріклітинного цГМФ і закриття натрієвих каналів, зменшує вміст у фоторецепторі не тільки Na+, але і Ca2+. В результаті пониження концентрації Ca2+ активується фермент гуанилатциклаза, що призводить до синтезу цГМФ, і в клітині збільшується вміст цГМФ. Це приводить до гальмування функцій активованою світлом фосфодиестерази. Обидва цих процеси сприяють підвищенню вмісту цГМФ і гальмуванню активності фосфодиестерази, що повертає фоторецептор до початкового стану відкриття натрієвих каналів.
У сітківці на 100 млн. паличок і 3 млн. колбочок доводиться близько 1, 6 млн. гангліозних клітин. В середньому на одну гангліозну клітку конвергує 60 паличок і 2 колбочки. Існують великі відмінності між периферичними і центральними відділами сітківки в кількості паличок і колбочок, конвергуючих на гангліозні нейрони.
В області центральної ямки залишаються тільки колбочки (близько 35000), і кількість волокон зорового нерва, що виходять з цієї області, дорівнює кількості колбочок. Це створює високий ступінь гостроти зору в центральній ямці в порівнянні з щодо слабкою гостротою зору на периферії сітківки.
Нейрони сітківки синтезують L-глутамінову кислоту, гліцин, ГАМК, ацетилхолін. Деякі нейрони містять, його аналоги (індоламіни) і нейропептиди. Палички і колбочки в синапсах з біполярними клітками секретують глутамат. Різні амакринні клітини виділяють, гліцин, дофамін, ацетілхолін і індоламін, завдяки чому можливе збудження і гальмуанния, що забезпечує обробку інформації.
Завдання 1.: Намалювати схему зорових шляхів та проаналізувати рівні обробки інформації.
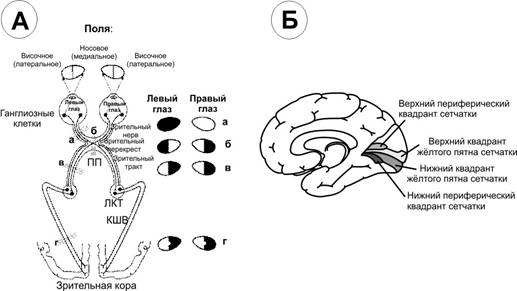
Зорові шляхи (А) і центри кори (Б). Літерами показано місце пошкодження зорових шляхів і зміни поля зору.
У колонках зорової кори є нейрони, виконуючі абсолютно певні функції (наприклад, аналіз контрасту, меж і напрямів ліній зорового образу й ін.).
У основі детекторної функції нейронів лежить, перш за все, аналіз контрасту зорового образу. Спочатку виявляються межі, ступінь і градієнт контрасту, після чого інші нейрони деталізують зоровий образ об'єкту. Розташовані в зовнішніх шарах зорової кори нейрони є детекторами ліній, певної довжини, кутів і інших форм зорових образів. Деякі групи нейронів визначають межі ліній і їх напрям (детектори орієнтацій). Це прості клітини. Інші клітини визначають напрям ліній при їх вертикальному або бічному зсуві (комплексні клітини).
При дослідженні кольорового зору слід проаналізувати механізми сприйняття кольору.
Сприйняття кольору - це функція колбочок. Існує три типи колбочок, кожний з яких містить тільки один з трьох різних зорових пігментів (червоний, зелений і синій).
Трихромазія - можливість розрізняти будь-які кольори, визначається присутністю в сітківці всіх трьох зорових пігментів (для червоного, зеленого і синього - первинні кольори). Ці основи теорії кольорового зору запропонував Томас Юнг у 1802р. і розвинув Г.Гельмгольц.
Д іхромазія (колірна сліпота, або дальтонізм) — дефекти колірного сприйняття (переважно у чоловіків; наприклад, в Європі різні спадкові дефекти у чоловіків складають 8% загальній популяції) по одному з первинних кольорів. Діхромазію підрозділяють на протанопію, дейтанопію і тританопію (від грецького - перший, другий і третій (є на увазі порядкові номери первинних кольорів: відповідно червоний, зелений, синій).
Повна колірна сліпота - ахромазія зустрічається украй рідко. При ахромазії вражається весь колбочковий апарат, і людина бачить всі предмети в різних відтінках сірого.
Формування гіперполяризаційного рецепторного потенціалу у колбочках при дії певного виду світлового спектру здійснюється так само, як і у паличках.
Таким чином, комплексне дослідження зорових функцій дозволяє оцінити стан зорової системи.
Дослідження слухової сенсорної системи.
Дослідження ролі допоміжних структур у дослідах Вебера і Рінне дозволяє оцінити стан допоміжних структур слухової сенсорної системи – повітряну і кісткову провідність до слухових рецепторів Кортиєва органа... Після цього слід пояснити механізми активації слухових рецепторів Кортиєва органа.
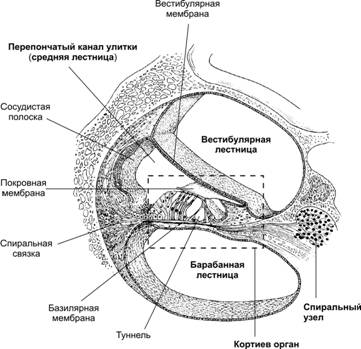
Схема каналів внутрішнього вуха і Кортиєва органа, розташованого на базілярній мембрані.
Слухові рецептори є вторинними механорецепторами і розташовані на основній (базілярній) мембрані завитка. Волосини рецепторів Кортиєва органа торкаються покривної мембрани.
Звукові хвилі викликають коливання барабанної перетинки, слухових кісточок, останні викликають коливання перилімфи, яка бере початок від овального вікна верхньої вестибулярної сходини і через гелікотрему сполучається з перилімфою нижньою базальної сходини, остання закрита мембраною круглого вікна.
Коливання перилімфи верхньої вестибулярної сходини передається через вестибулярну мембрану на ендолімфу середньої сходини, що призводить до коливання базілярної мембрани, яка є еластичнішою за покрівну мембрану. При коливанні базілярної мембрани волосини слухових рецепторів кортиєва органа торкаються покривної мембрани, що призводить до зменшення проникності мембрани рецептора до іонів калію і виникнення деполяризації, рух в протилежному напрямку призводить до гіперполяризації.
Інформація передається на аферентні нерви (УІІІ пара), тіла цих нейронів розташовані у спіральному ганглії і далі по слуховим провідним шляхам аж до слухової кори.
Людина сприймає звуки в діапазоні коливання від 16 до 20000 Гц. Розрізняти звуки високої і низької частоти дозволяє активація рецепторів, що розташовані на базілярній мембрані. Рецептори, що розташовані близько до овального вікна на базілярній мембрані в основі завитка, активуються звуками високої частоти, а звуки низької частоти викликають коливання базілярної мембрани і активацію слухових рецепторі, що розташовані ближче до гелікотреми.
Тому зниження гостроти слуху на аудиіограмах до високих чи низьких частот дозволяє студентам визначити механізми кодування інформації у слуховій сенсорній системи залежно від розташування слухових рецепторів, які активуються.
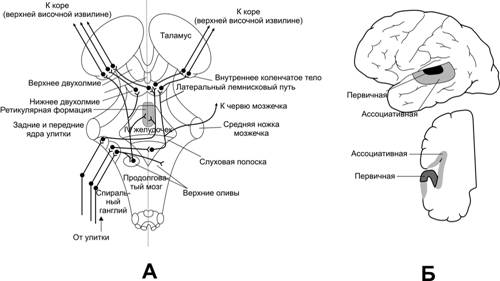
А.Схема основних слухових шляхів та рівнів обробки інформації. Б.Слухова кора.
Проекційні області слухової кори розташовуються не тільки у верхній частині верхньої скроневої звивини, але і тягнуться на зовнішню сторону скроневої частки, захоплюючи частину острівкової кори і тім'яної покришки.
Первинна слухова кора безпосередньо отримує сигнали від внутрішнього (медіального) колінчастого тіла, тоді як слухова асоціативна область повторно збуджується імпульсами з первинної слухової кори і таламічних областей, що граничать з медіальним колінчастим тілом.
Напрям джерела звуку. Два вуха, що працюють в унісон, можуть виявляти джерело звуку по різниці в гучності і часі, який йому потрібний, щоб досягти обох сторін голови. Чоловік визначає звук, що йде до нього, двома шляхами.
1)Часом затримки між надходженням звуку в одне вухо і в протилежне вухо. Спочатку звук поступає до вуха, що знаходиться ближче до джерела звуку. Звуки низької частоти огинають голову через їх значну довжину. Якщо джерело звуку знаходиться по середній лінії спереду або ззаду, то навіть мінімальне зрушення від середньої лінії сприймається людиною. Таке тонке порівняння мінімальної різниці в часі приходу звуку здійснюється ЦНС в очкках, де здійснюється конвергенція слухових сигналів. Цими точками конвергенції є верхня олива, нижні двогорбики, первинна слухова кора.
2)Відмінністю між інтенсивністю звуків в двох вухах. При високих частотах звуку розмір голови помітно перевищує довжину звукової хвилі, і хвиля відбивається головою. Це приводить до виникнення різниці в інтенсивності звуків, що приходять до правого і лівого вуха.
|
|