Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Тема: Физиологические мышечной ткани.
|
|
Время 90 минут
Учебные и воспитательные цели:
Изучить классификацию, значение мышц, особенности скелетных и мышц внутренних органов.
ЛИТЕРАТУРА
Основы физиологии человека. Под редакцией Б.И.Ткаченко. - С.-Петербург, 1994. - Т.1. - С. 146 - 165.
Физиология человека. Под редакцией Р.Шмидта и Г. Тевса. - М., Мир.- 1996. - Т.1..
- Физиология человека. Под ред. В.М.Покровского, Г.Ф. Коротько.М., Медицина. – 2000.-Т..1- С 71 – 93.
МАТЕРИАЛЬНОЕ ОБЕСПЕЧНИЕ
1. Мультимедийная презентация 26 слайдов.
РАСЧЕТ УЧЕБНОГО ВРЕМЕНИ
| № п/п | Перечень учебных вопросов | Количество выделяемого времени в минутах |
| 1. | Классификация и функции мышц. | |
| Теория мышечного сокращения. | ||
| Режимы и виды сокращения мышц. | ||
| Сила мышц. | ||
| Утомление мышц. | ||
| Физиологические особенности гладких мышц. |
Всего 90 мин
1. Классификация и функции мышц. Виды мышечных тканей.
МЫШЕЧНЫЕ ТКАНИ - ткани различного происхождения и строения, для которых ведущей функцией является сократимость, а основными органеллами - миофибриллы. Все тканевые элементы имеют удлиненную форму.
Выделяют три вида мышечных тканей:
1. Поперечно-полосатую (скелетную) мышечную ткань.
Является произвольной возбудимой тканью. Формирует скелетные мышцы, мышцы ротовой полости, верхней трети глотки, наружные сфинктеры мочеиспускательного канала и прямой кишки. Тканевыми элементами служат мышечные волокна - симпласты, которые содержат 4 элемента, обеспечивающих сократительную функцию мышц:
а) миофибриллы, образованные параллельно ориентированными нитями актина и миозина, соединенные концами друг с другом;
б) тропонин-тропомиозин регулирующий комплекс (управляет актом сокращения);
в) саркоплазматический ретикулум, образующий систему T-трубочек и L-каналов, которые содержат ионизированный кальций, запускающий сокращение;
г) систему энергетического обеспечения (митохондриальные комплексы).
2. Гладкомышечную ткань.
Непроизвольно возбудимая ткань, формирует мышечные оболочки внутренних органов, все внутренние сфинктеры, входит в состав стенки сосудов. Тканевым элементом является гладкомышечная клетка - миоцит веретеновидной или звездчатой формы. Сократительные элементы представлены:
а) миозиновыми нитями, проходящими вдоль оси клетки и актиновыми нитями, образующими 3-мерную сеть;
б) слабо развитым тропонин-тропомиозиновым комплексом;
в) не развитым эндоплазматическим ретикулумом, (необходимый для инициирования мышечного сокращения ионизированный кальций поступает из внешней среды клетки);
г) системой энергетического обеспечения.
3. Сердечную мышечную ткань (миокард). Сочетает в себе черты скелетной и гладкомышечной ткани. Непроизвольно возбудимая ткань, обладающая собственной ритмической активностью. Тканевым элементом служат вытянутые клетки - кардиомиоциты, которые при помощи нексусов (электронноплотных контактов) объединены в функциональные волокна. Среди кардиомиоцитов есть атипичные клетки, образующие проводящую систему. Сократительные элементы клетки представлены:
а) миофибриллярными комплексами, как у скелетных мышц;
б) развитым тропонин-тропомиозиновым комплексом;
в) сочетанным поступлением кальция как из внешней среды, так и из саркоплазматического ретикулума;
г) системой энергетического обеспечения.
Скелетные мышцы обеспечивают сохранение позы, взаимное расположение частей тела и перемещение тела в пространстве, дыхание, прием пищи. Благодаря сокращениям сердечной мышцы, осуществляется циркуляция крови по сердечно-сосудистой системе. Мышцы внутренних органов обеспечивают перистальтику желудка, кишечника, мочеточника, маточной трубы, тонус кровеносных и лимфатических сосудов, опорожнение кишечника, мочевого пузыря.
Двигательной единицей скелетной мышцы является мотонейрон ЦНС и иннервируемые им мышечные волокна.
В функциональном отношении различают фазные мышечные волокна (обеспечивают движения, связанные с перемещением тела в пространстве) и тонические мышечные волокна (обеспечивают длительно протекающие сократительные процессы, например, сохранение позы). В фазных волокнах генерируется потенциал действия, который и распространяется по мембране. В тонических волокнах возбуждение возникает градуально в зависимости от силы стимула. Фазные мышечные волокна в свою очередь подразделяются на быстрые и медленные мышечные волокна.
| Быстрые волокна | Медленные волокна | |
| Длительность сокращения | 10 - 30 мс | 100 и более |
| Тетанус развивается при частоте стимуляции (им/сек) | 10 - 15 | Около 50 |
| Сила сокращения | Больше | Меньше |
| Утомление | Развивается быстрее | Развивается медленнее |
2. Строение поперечно-полосатой мышцы.
Скелетные мышцы позвоночных состоят из клеток (мышечных волокон). Каждое мышечное волокно - это многоядерная клетка цилиндрической формы диаметром 20 - 80 мкм и длиной от нескольких миллиметров до десятков сантиметров, соответствующей длине мышцы. Многоядерные волокна исчерченных мышц образуются путём слияния одноядерных клеток - миобластов. В мышце волокна объединены в пучки по 20 - 40 волокон и отделены друг от друга соединительной тканью. К этой оболочке со стороны мышечной клетки прилегает плазматическая мембрана - сарколемма. В каждом волокне имеется около 2000 миофибрилл диаметром 1-2 мкм, проходящих от одного конца волокна до другого. Особенностью их строения является поперечная исчерченность. Каждая миофибрилла разделена с помощью Z - дисков на саркомеры длиной 2 - 3 мкм. Каждый саркомер состоит из перекрывающихся толстых протофибрилл (миозиновых) длиной 1, 6 мкм и диаметром 16 нм и тонких протофибрилл (актиновых) длиной 1 мкм и диаметром 8 нм. Тонкие протофибриллы прикрепляются к Z-дискам и образуют изотропную I-полосу. Толстые протофибриллы в центре саркомера образуют анизотропную А-полосу.
В области перекрытия протофибриллы образуют гексогональную структуру, в которой одну миозиновую нить окружают шесть актиновых. Взаимодействие между толстыми и тонкими протофибриллами осуществляется с помощью поперечных мостиков, которые выступают из миозиновых протофибрилл.
Основным белком тонкой протофибриллы является актин в глобулярной (сжатой, cкрученной) форме - G - АКТИН (молекулярная масса 42000 Д). Из глобул G- актина образуются путём полимеризации нити F-актина. Нити F-актина образуют двухтяжевую структуру, состоящую из двух скрученных нитей с периодом 73 - 74 нм. Кроме актина, тонкая протофибрилла содержит фибриллярный белок тропомиозин (ТМ) молекулярной массой = 68 000 Д и глобулярный белок тропонин (ТН) молекулярной массой = 70 000 Д. Три белка в тонкой протофибрилле находятся в определённом соотношении: на 7 глобул двойной нити актина приходится по одной молекуле тропомиозина и тропонина. Стержнеподобные молекулы тропомиозина длиной около 40 нм располагаются в бороздках актиновой суперспирали и взаимодействуют конец к концу. С каждой молекулой тропомиозина связывается молекула тропонина, состоящая из трёх субъединиц: тропонин - связывающей субъединицы - (ТН-Т, М.м = 30500 Д), ингибиторной субъединицы - (ТН-I, М.м. = 20700 Д) и кальций-связывающей субъединицы - (ТН- С, М.м. = 17850 Д). ТН - С имеет четыре кальцийсвязывающих центра. АТФ-азная активность миозина открыта В.А.Энгельгардтом и М.Н.Любимовой в 1939 г. и является главной функциональной характеристикой миозина. В процессе гидролиза АТФ миозином освобождается энергия, обеспечивающая движение головок миозина.
3. Теория мышечного сокращения.
Предложена А.Хаксли (1957). В основе мышечного сокращения лежит взаимодействие между толстой и тонкой протофибриллами, регулируемое ионами кальция. Ионы кальция, связываясь с тропонином С, вызывают ряд изменений в структуре и организации белков тонкой протофибриллы, которые приводят к взаимодействию молекул актина и миозина. Тонкие и толстые протофибриллы способны смещаться по отношению друг к другу. В неактивном состоянии мышцы, когда концентрация ионов кальция ниже 1´ 10-8 М, тропомиозин ориентируется под углом 50о по отношению к центру тонкой протофибриллы, в результате чего ТН-I ингибирует взаимодействие головки миозина с актиновой протофибриллой. При возбуждении мышцы ионы кальция связываются с ТН-С, изменяется положение тропомиозина, головка миозина взаимодействует с тонкой протофибриллой, развивая активную тянущую силу.
4. Электромеханическое сопряжение
Связь между возбуждением и сокращением мышечного волокна описана А.Хаксли (1959). Осуществляется при помощи системы поперечных трубочек поверхностной мембраны (Т-системы) и внутриволоконного саркоплазматического ретикулума. Деполяризация, вызываемая потенциалом действия, распространяется на Т - систему и стимулирует освобождение ионов кальция из полостей ретикулума. Взаимодействие ионов кальция с регуляторным белком тропонином С приводит к активации системы сократительных белков актина и миозина. Механизм генерации потенциала действия принципиально не отличается от этого процесса в нейроне. Скорость его распространения по мембране мышечного волокна 3 - 5 м/c.
5. Режимы и виды сокращения мышц
Режимы сокращения мышцы: изотонический (когда мышца укорачивается при неизменном внутреннем напряжении, например, при нулевой массе поднимаемого груза) и изометрический (при этом режиме мышца не укорачивается, а лишь развивает внутреннее напряжение, что бывает при нагрузке неподъёмным грузом). Ауксотонический режим - при сокращении мышцы с нагрузкой вначале в мышце возрастает напряжение без укорочения (изометрический режим), затем, когда напряжение преодолевает массу поднимаемого груза, укорочение мышцы происходит без дальнейшего роста напряжения (изотонический режим).
Различают виды сокращений: одиночное и тетаническое. Одиночное сокращение возникает при действии на мышцу одиночного нервного импульса или однократного толчка тока. В миоплазме мышцы происходит кратковременный подъём концентрации кальция, сопровождаемый кратковременной работой - тягой миозиновых мостиков, сменяющейся покоем. В изометрическом режиме одиночное напряжение начинается через 2 мс после развития потенциала действия, причём напряжению предшествует кратковременное и незначительное латентное расслабление.
Тетанус - это сложное сокращение, возникающее при стимуляции с частотой выше, чем длительность одиночного мышечного сокращения. Тетанус бывает зубчатый, если мышца совершает незначительные колебания на высоте амплитуды сокращения, и гладкий - при постоянном во времени сокращении. При относительно малой частоте раздражений возникает зубчатый тетанус, при большой частоте - гладкий тетанус. Чем быстрее сокращаются и расслабляются волокна мышцы, тем чаще должны быть раздражения, чтобы вызвать тетанус.
В естественных условиях мышечные волокна работают в режиме одиночного сокращения только тогда, когда длительность интервала между разрядами мотонейронов равна или превышает длительность одиночного сокращения иннервируемых данным мотонейроном мышечных волокон. В режиме одиночного сокращения мышца способна работать длительное время без утомления, совершая при этом минимальную работу. При увеличении частоты разрядов развивается тетаническое сокращение. При зубчатом тетанусе происходит непрерывное нарастание силы сокращения и выполняемой работы. Во время гладкого тетануса мышечное напряжение не изменяется, а поддерживается на достигнутом уровне. В таком режиме мышца человека работает при развитии максимальных изометрических усилий. Работа мышцы (А) измеряется произведением массы груза (Р) и расстояния (H), на которое этот груз перемещается. 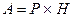
Работа может быть динамической (преобладают изотонические режимы сокращения) или статической. Она может быть преодолевающей и уступающей.
Расслабление мышцы.
Восстановление потенциала покоя мембраны прекращает поступление из саркоплазматического ретикулума ионов кальция и дальнейший сократительный процесс. Кальций в миоплазме активирует Са-АТФ-азу, кальциевый насос осуществляет активный перенос этого иона в саркоплазматический ретикулум. Возврат мышцы в исходное, растянутое положение определяется массой костей скелета, связанных с данными мышцами и создающими растягивающее усилие после прекращения процесса сокращения. Вторым моментом является упругость мышцы, которая преодолевается в момент сокращения. Структурной основой упругости мышцы являются:
Поперечные мостики.
Z - диски.
Участки прикрепления концов миофибрилл к сухожильным элементам мышечного волокна.
Наружные соединительнотканные элементы мышцы и её волокна.
Места прикрепления мышц к костям.
Продольная система саркоплазматического ретикулума.
Сарколемма мышечного волокна.
Капиллярная сосудистая сеть мышцы.
6. Сила мышц
Сила мышц измеряется максимальным грузом, который она способна поднять или максимальным напряжением, которое она способна развить в условиях изометрического сокращения. Сила мышц пропорциональна площади поперечного сечения (количеству мышечных клеток). Физиологическое поперечное сечение проводится перпендикулярно мышечным волокнам. Поэтому мышцы с косым расположением мышечных волокон обладают большей силой. Абсолютная мышечная сила определяется делением максимального груза, который мышца способна поднять, на площадь физиологического поперечного сечения.
7. Утомление мышц
Утомлением мышцы называется снижение работоспособности мышцы в процессе выполнения мышечной работы. Причиной снижения работоспособности (утомления) является уменьшение в мышечной клетке запасов питательных веществ, накопление метаболитов, истощение медиатора в нервно-мышечном синапсе.
Роль тренировки
Тренировка - система физических упражнений и режим их реализации - имеет целью увеличение долговременности и объема выполняемой мышечной работы. Это достигается за счет увеличения в организме массы мышечной ткани, перестройки обмена веществ в мышце, изменения в системах кровоснабжения мышц, дыхания, нервной регуляции мышечной деятельности. Метаболические изменения в мышце позволяют выполнять работу в условиях сниженного поступления к мышце кислорода (в анаэробных условиях), увеличиваются кислородная емкость крови, минутный объем сердечного выброса, минутный объем дыхания, скорость кровотока.
8. Физиологические особенности гладких мышц.
Гладкомышечные клетки делят на два типа: сосудистого и висцерального. Гладкомышечная клетка сосудистого типа имеет размеры 20´ 5-10 мкм, у клеток висцерального типа эти параметры равны 100-200´ 10-20 мкм. Плазматическая мембрана гладкомышечной клетки имеет особые впячивания - кавеолы, благодаря которым площадь мембраны увеличивается на 70%. Между рядами кавеол располагаются электронно-плотные тяжи, связанные с актиновыми протофибриллами. Снаружи плазматическая мембрана покрыта базальной мембраной и отделена от нее электронно-светлой областью. Комплекс базальной и плазматической мембраны называют сарколеммой. Ядро располагается в средней утолщенной части клетки и не имеет особенностей по сравнению с клетками других типов. СПР состоит из гранулярной и гладкой части, гранулярная часть лежит у ядра, гладкая - вблизи плазмалеммы и часто тесно контактирует с кавеолами. Гладкомышечные клетки лежат в виде параллельно ориентированных пучков, расстояние между ними заполнено коллагеновыми и эластическими волокнами, фибробластами, питающими магистралями. Между клетками имеются тесные контакты: 1) нексусы - щели ~2-3 нм, имеющие ионные каналы и 2) десмосомоподобные контакты - щели 20-60 нм, заполненные электронно-плотным веществом, переходящим в базальную мембрану, выполняют механическую функцию, связывая клетки.
Основу сократительного аппарата составляют миозиновые и актиновые протофибриллы. В ГМК актиновых протофибрилл намного больше, чем в поперечно-полосатом мышечном волокне. Соотношение актин/миозин = 5: 1.
Миозиновые протофибриллы имеют толщину 14 нм, длину 2, 6 мкм - т.е. заметно больше, чем в скелетных мышцах, и состоят из миозиновых молекул. В миоците миозин агрегирует в толстые протофибриллы, с образованием той же пространственной структуры, как и миозин скелетных мышц.
Актиновые протофибриллы имеют удлиненную форму, гексагональны на срезе и представляют собой агрегаты актина, тропомиозина и кальдесмона. Глобулы G-актина полимеризуются в 2 цепи F-актина, образующие видоизмененный остов молекулы фибриллы. Тропомиозина в ГМК в 7 раз меньше, чем в скелетных мышцах, они расположены в желобках каркаса актиновой глобулы, контактируя с 7 глобулами F-актина. Тропонин скелетных мышц в ГМК отсутствует.
Помимо описанных 2 типов протофибрилл в ГМК есть промежуточные фибриллы, весьма стойкие к различным денатурирующим агентам. Эти протофибриллы образуют трехмерную сеть, крепящуюся к плотным тельцам десмосом и межкавеолярных тяжей, которая служит сократительным остовом (своеобразной внутриклеточной «точкой опоры», к которой и прилагается сила сокращения).
В ГМК протофибриллы, в отличие от скелетной мышцы, не пакуются в миофибриллы, саркомера как такового нет, роль Z-пластинок выполняют плотные тельца. В ГМК отсутствует Ca++-тропониновая система активации сокращения, вместо нее ведущая роль принадлежит системе фосфорилирования. Возрастание концентрации внутриклеточного кальция [Cai++] и взаимодействие его с кальмодулином приводит к повышению аффинности (сродства) комплекса Са++-кальмодулин к киназе L-цепи миозина (ее каталитической единицы). Происходящее при этом фосфорилирование легкой цепи миозина вызывает активацию актинзависимой Mg++-АТФ-азы миозина и генерацию сокращения. Ингибиторами фосфорилирования L-цепи миозина служат цАМФ-зависимые протеинкиназы, которые в отсутствие Са++ фосфорилируют 2 центра киназы L-цепей и снижают тем самым их аффинность к комплексу Са++-кальмодулин, а это, в конечном итоге, уменьшает фосфорилирование L-цепей. Помимо фосфорилирующей системы сокращение ГМК регулирует независимый Са++-чувствительный механизм. При этом фосфорилирующая система контролирует скорость циклирования поперечных мостиков (присоединения-отсоединения миозина к актину) в фазу развития напряжения, а Са++-чувствительный механизм - в фазу тонуса.
Электрофизиологические явления в ГМК. Если в анатомическом смысле гладкомышечная ткань представляет собой слаженный ансамбль отдельных клеток, то в функциональном плане - это электрический синцитий. ГМК имеет ряд электрофизиологических особенностей: 1) клетка не способна (за небольшим исключением) генерировать «полноценный» потенциал действия (ПД), вместо этого на мембране возникает локальная деполяризация; 2) электротонические потенциалы способны распространяться на большие расстояния, не затухая. Распространение электротонического потенциала идет по нексусам, обладающих чрезвычайно низким сопротивлением.
Потенциал покоя (ПП) ГМК. Проводимость мембраны ГМК для одновалентных ионов составляет pK+: pNa+: pCl- = 1: 0, 2: 0, 6-0, 7. Однако, в генерации ПП клетки играют ведущую роль не только эти ионные токи, но и электрогенный компонент Na+-K+ насоса (что в меньшей степени характерно для ПП скелетного мышечного волокна). ПП клетки колеблется в пределах -50¸ -70 мВ, тогда как пороговый потенциал (при котором включаются потенциалзависимые Са++ каналы) составляет -30¸ -35 мВ. Основной вклад в ПП ГМК вносят ионы К+ (Ko+= 2-3 мМ, Кi+= 160 мМ). Ионы Na+ и Cl- оказывают на величину ПП клетки лишь мягкое «регулирующее» действие.
Потенциал действия ГМК. Спонтанная активность ГМК проявляется в виде медленной волны деполяризации (15-20 мВ, длительность 250-400 мс) и предшествующего ей пикового потенциала (25-50 мВ, длительность 50 мс). Часто вслед за первым идет второй пиковый потенциал, чем больше пауза после первого пикового потенциала, тем больше амплитуда и меньше длительность второго пика. Второй пиковый потенциал в отличие от первого никогда не прерывает развитие медленной волны деполяризации. Спонтанная активность ГМК имеет миогенную природу, за счет пейсмекерной активности ГМК, но в целом зависит от уровня ПП (чем он ниже - тем меньше активность). Ионная природа спонтанной активности обусловлена изменением концентрации ионов Са++ у наружной стороны мембраны. Удаление Са++ вначале усиливает спонтанную активность, а затем угнетает пиковый, и далее, медленный ее компонент. Увеличение Сао++ сопровождается снижением частоты спонтанных разрядов и увеличением их амплитуды. Ионы Са++ обеспечивают фазу деполяризации ПД и медленной волны, тогда как ионы К+ и выходящий К+ ток - фазу реполяризации. ПД ГМК артериол зависит от мощности их симпатической иннервации, чем она выше, тем больше ПД генерируется.
Роль Са++ в сопряжении возбуждения и сокращения. В отличие от скелетных мышц, где функцию сопрягающего агента между мембраной и актомиозиновым комплексом выполняет Т-система и СПР, в ГМК эти образования либо отсутствуют (Т-система), либо их недостаточно (СПР составляет примерно 2-7% объема цитоплазмы). В ГМК функцию сопряжения выполняют, очевидно, кавеолы плазматической мембраны ГМК и приближенные к ним участки СПР.
Важную роль в сопряжении процесса возбуждения и сокращения играют ионы Са++, которые снимают тормозящее воздействие тропонина на актин-миозиновое взаимодействие и запускают сокращения. В ГМК исходная концентрация Са++ составляет 2, 0-2, 8´ 10-7 моль/л, повышаясь до 3, 8´ 10-6 моль/л на максимуме сокращения. Источниками Са++ для ГМК служат: 1) ионизированный Са++ внутриклеточной среды; 2) Са++ токи плазматической мембраны через 2 класса Са++-каналов и Na+-неселективные каналы; 3) внутриклеточные депо Са++ - митохондрии и СПР, внутренняя поверхность плазматической мембраны с ее Са++-связывающими белками. В отличие от скелетной мышцы, в ГМК ведущая роль принадлежит не СПР, а плазмалеммальным токам, т.к. благодаря относительно малому объему каждой клетки по сравнению с поперечно-полосатым мышечным волокном диффузионное расстояние плазмалемма - миофибрилла падает.
ЗАКЛЮЧЕНИЕ
Ведущей функцией мышечных тканей является сократимость. Скелетные мышцы организма обеспечивают сохранение позы, взаимное расположение частей тела и активное перемещение тела в пространстве, дыхание. Это делает жизнь свободной, не ограниченной в пространстве, позволяет организму активно взаимодействовать с окружающей средой, преобразовывать ее. Благодаря сокращениям сердечной мышцы, осуществляется циркуляция крови по сердечно-сосудистой системе. Мышцы внутренних органов обеспечивают перистальтику желудка, кишечника, мочеточника, маточной трубы, тонус кровеносных и лимфатических сосудов, опорожнение кишечника, мочевого пузыря.
Учебно-контрольные вопросы по теме лекции
1. Одиночное мышечное сокращение и тетанус. Влияние силы раздражителя на величину мышечного сокращения. Работа мышц.
2. Микроскопическое строение исчерченных (поперечно-полосатых) мышц. Механизм мышечного сокращения.
3. Функциональные особенности гладких мышц.
4. Утомление мышц, теории утомления. Влияние симпатической нервной системы на утомление в организме. Утомление в целом организме. Значение тренировки, активный отдых.
|
|