Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Становление системы биогеохимических циклов
|
|
Г. А. Заварзин
©2003г.
Институт микробиологии РАН
Поступила в редакцию 28.12.2002 г.
Принята к печати 21.04.2003 г.
ПАЛЕОНТОЛОГИЧЕСКИЙ ЖУРНАЛ, 2003, № 6, с. 16-24
В результате открытий последних десятилетий, которые можно отнести к бактериальной палеонтологии в широком смысле слова, представления об эволюции геосферно-биосферной системы дали основу новому мировоззрению, в котором большее значение придается кооперативным взаимоотношениям в рамках большой системы. Рассмотренная система находится вне области традиционных для биологов популяционных пределов, составляющих основу дарвинизма. Поэтому область геосферно-биосферных систем удобно обозначить как внедарвинистскую. В ней наряду с определяющими прямыми связями от геосферы к биоте большую роль играют трансформирующие обратные связи. Прокариотное сообщество, катализирующее систему биогеохимических циклов, служит основой для дальнейшей эволюции. Возрастающая сложность живых организмов не упраздняет ключевую роль " невидимых" для существования биосферы, а накладывается на созданную ими систему. Эволюция в рассматриваемом масштабе носит выраженный аддитивный характер.
История геосферно-биосферных систем. История Земли прослеживается палеонтологически для поверхности суши от современности с доминирующим растительно-почвенным покровом, кончая силуром, когда этого покрова еще не было, а в море пищевая пирамида с завершающими ее животными с зоотрофным питанием прослеживается до начала кембрия. Венд составляет эпоху многоклеточных водорослей и паразоа с преимущественно фаготрофным питанием; население на суше в этот период неясно. Ниже идет эпоха микробной жизни протерозоя, в котором биолог может выделить два этапа: один, с доминированием эвкариотных протист, как в современном океане, и другой вплоть до начала геологической летописи с исключительно прокариот-ными организмами. Переход между этими этапами располагается около 1 млрд. лет назад и относится к началу неопротерозоя, хотя первые находки эвкариот относятся к более раннему периоду. Условно эти этапы можно назвать " проти-стий" для эпохи распространения протист и " про-кариотий" для первоначального бактериального этапа с микрофоссилиями возрастом 3.5 млрд. лет. Эти микрофоссилии относятся к находкам в формациях Барбертон (Южная Африка) и Пил-бара (Австралия), в то время как находки в железистых кварцитах Иссуа (3.8 млрд. лет) поставлены под сомнение (Westall et al., 2001). Ниже располагается эпоха преимущественно геологических процессов становления Земли и формирования ее геологических оболочек.
Моделью для реконструкции прокариотной биосферы служат экосистемы реликтовых микробных сообществ, являющихся аналогами микробных сообществ прошлого и сохраняющихся преимущественно в экстремальных условиях, недоступных для эвкариот, единичные представители которых появляются там как вторичные вселенцы. Отсюда можно сделать заключение, что коридор предпочтительных мест обитания сокращается с увеличением сложности организмов. Реликтовые микробные сообщества составляют экспериментальную базу для актуалистической бактериальной палеонтологии. При этом следует иметь в виду, что актуалистический подход, исходящий из персистентности своих объектов, в принципе не допускает датировки; использовать аналогию реликтовых сообществ для реконструкции определенной эпохи можно лишь на основе геологических фактов, допускающих такую реконструкцию. Однако в отличие от палеонтологии высших организмов в бактериологии можно основываться не на отдельных " живых ископаемых", а на целых сообществах и реконструировать потоки вещества в них на основе трофических сетей. Тесная связь бактерий с геосферой позволяет использовать геохимические продукты их жизнедеятельности для определения их функциональных свойств, которые нельзя или трудно установить на основе чисто палеонтологических данных по морфологии микрофоссилии. Это позволяет с большой вероятностью реконструировать биогеохимические циклы прошлого.
Становление системы биогеохимических циклов имело место в период господства на Земле бактерий и может быть понято на основании сопоставления современных процессов взаимодействия бактериальных сообществ с геологическими факторами. Последовательность событий с очевидностью указывает на то, что формирование предыдущего этапа служит фундаментом, базисом для надстройки на него следующего и вся система представляет собой пирамиду, в которой сохраняемое прошлое служит основой и условием настоящего. Данные бактериальной палеонтологии убеждают, что массовое распространение эукариот-протист произошло в течение неопротерозойской революции, которая совершалась около 1 млрд. лет назад. До этого господствовала прокариотная биосфера, которая и явилась основой развития жизни в процессе аддитивной эволюции, включающей прошлое как основу настоящего.
При анализе эволюции геосферно-биосферной системы необходимо различать прямые и обратные связи. Происходит ли приспособление микробного мира к задаваемым извне геосферными силами условиям или же биота создает свою среду обитания, используя неорганическую материю лишь как материал?
Анти-Гея. За рубежом большое распространение получила гипотеза Геи, согласно которой Земля представляет собой один огромный организм, регулирующий обстановку для оптимального развития жизни (Lovelock, 1995). Кстати, впервые метафора " Земля как огромный организм", оперирующий на основе катализа реакций микробами - " ферментами", как их называли в то время, - была высказана автором открытия хемосинтеза С.Н. Виноградским в 1896 г. (Виноград-ский, 1996), задолго до создания представления о биосфере. Наибольшее значение в концепции Геи имеет регуляция атмосферы как в отношении состава газов, так и изменения альбедо. По моему мнению, гипотеза Геи представляет гипертрофию обратных связей в системе, получившую поддержку из-за придания чрезмерного значения парниковому эффекту. Кто-то назвал международную геосферно-биосферную программу IGBP -" глобальной биосферной программой". Это соответствует идущему от Вернадского пониманию биосферы как всей области, где встречается жизнь, вне зависимости от того, какую роль играют там процессы, обусловленные жизнью. Для последующего изложения будет удобнее разделить геосферу и биосферу, понимая под последней только ту область, где явления жизни играют определяющую роль. Роль обратных связей от биоты велика и приложима к временам негеологического масштаба, т.е. нескольких сотен тысяч лет, когда состояние геосферы можно считать постоянным. В геологическом масштабе времени приходится оперировать временами более 104 лет. В задачу нашего обзора входит рассмотрение процессов длительностью более сотен миллионов лет. При таком масштабе на первый план выходят проблемы, связанные с астрономическими факторами, какими являются светимость Солнца, положение планеты, но в первую очередь, с геологическими факторами. Именно геологические факторы играют роль прямых связей в системе по отношению к биоте - сумме дискретных живых организмов, а не метафорическому термину " живое вещество", наложившему чуждый смысл на понятие, обязательно включающее биоразнообразие. Представление о создании биотой исходной среды обитания заведомо неточно, поскольку среда, создаваемая абиотическими факторами, изначально предшествует обитанию.
К важным прямым связям относится дегазация Земли, приведшая к формированию первоначальной атмосферы и гидросферы и закончившаяся до начала геологической летописи осадочных отложений. Эта часть истории Земли относится, по определению В.Е. Хаина, к " фантомной" (Ха-ин и др., 1997). Большинство моделей исходит из очень быстрой, в течение немногих сотен миллионов лет, дегазации планеты с выделением одного эквивалента океана за 100 млн. лет (Franck, Bounama, 1995) с последующим гораздо более медленным выделением газов пропорционально скорости роста океанической коры, т.е. около 70 миллиардных частей СО2 в год. О.Г. Сорохтин и С.Д. Ушаков (2002) допускают гораздо более позднюю дегазацию с ростом океана в 1-2 км3 в год. Поздняя дегазация ставит вопрос о возможности взаимодействия первичной микробиоты с эндогенными газами, а не только с газами рецикла, как это происходит сейчас на островных дугах (Заварзин и др., 1989). В гораздо большей степени к фантомной области относится вопрос происхождения жизни на Земле, находящийся в области произвольных гипотез. Поэтому при обсуждении становления биогеохимических циклов отсутствует стартовая точка и область ранней истории Земли приходится реконструировать сверху вниз, что является нормальным путем исследования и реконструкции на основе принципа " обратной истории". Применение актуалистичес-кого подхода и анализа явлений от следствия к причинам имеет свои ограничения и на них следует обратить внимание в первую очередь. Несмотря на неоднократные попытки, актуалистический подход вряд ли применим к раннему архею.
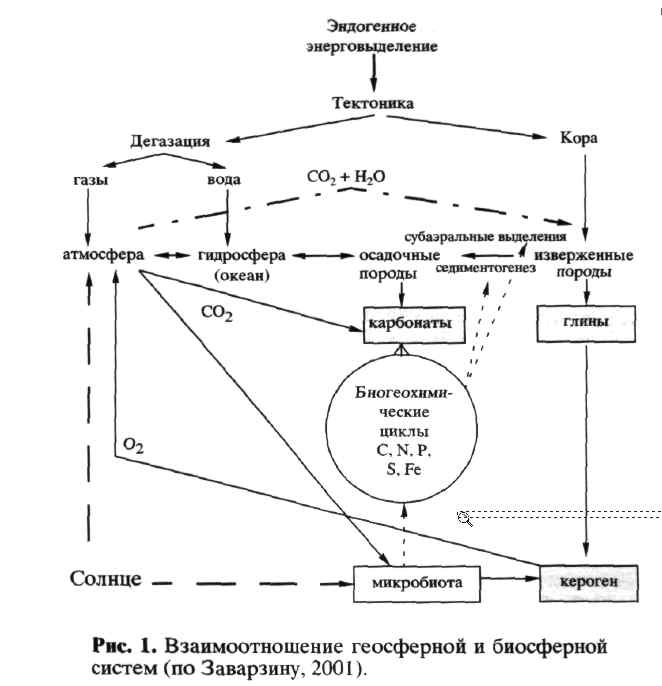
В геосферной системе можно выделить следующие основные блоки, соотношение которых было рассмотрено в связи с определяющими прямыми связями от геосферных факторов в гео-сферно-биосферной системе (Заварзин, 2001) (рис. 1; следует обратить внимание на различие преимущественных источников энергии - эндогенной для геосферы и солнечной для биосферы; в схеме выявляются два узла - атмосферы и химического выветривания). Движущей силой в этой системе служит атмосферный гидрологический цикл, в свою очередь зависящий от энергии Солнца и перераспределения воздушных масс по земному шару. Ключевым процессом служит субаэ-ральное химическое выветривание, ведущее к формированию коры выветривания за счет реакции СО2 с породами, конечным результатом которого служит образование обедненных катионами глин и соответствующих им новообразованных карбонатов Са и Mg. Реакция MeSiO3 + СО2 = = МеСО3 + SiO2 считается основной для освобождения атмосферы от углекислоты дегазации и перевода ее в карбонатные породы осадков.
Катионы в виде бикарбонатов с речным стоком выносятся в океан. Скорость абиогенных процессов углекислотного выветривания считается недостаточной для преобразования, и необходимо найти пути его ускорения (Schwartzman, 1999). Известно, что коры выветривания вплоть до раннего докембрия сходны с современными, что указывает на единство процессов в субаэ-ральной оболочке Земли. Для формирования химического состава океана дополнительно к речному стоку значение имеет циркуляция воды через тонкую океаническую кору. Предполагается, что особое значение при этом играют узкие зоны спрединга.
Важнейшим звеном в рассматриваемой схеме служит формирование состава атмосферы под действием микробиоты (Заварзин, 1984). Кроме прямой продукции газов, влияние микробиоты определяется ее каталитическим воздействием через биогеохимические циклы на субаэральное химическое выветривание, седиментогенез, образование карбонатов, образование керогена. Образование керогена обусловливается скоростью его захоронения в тонкодисперсных осадках (глинах) и соответственно не только скоростью продукции Сорг, но и скоростью седиментогенеза с ключевым этапом - переходом в анаэробные условия (Tyson, 1995). Кероген в рассматриваемом масштабе времени ответственен за баланс по кислороду и, следовательно, за формирование окси-ческой атмосферы. Область интересов бактериальной палеонтологии обозначена на рис. 1 заштрихованными блоками осадков.
Не следует представлять планету как нечто однородное. Это приложимо только к хорошо перемешанной атмосфере. Во все времена существовал зависимый от климата цикл карбонатов кальция с растворением СО2 в холодных водах и соответственно мобилизацией кальция в виде бикарбонатов. Перенос вод глубинными течениями в меридиональном направлении приводил к отложению карбоната кальция в теплом мелководном океане с возвратом 1/2СО2 в атмосферу. Отложение карбоната кальция в субаэральных условиях не допускало повышения карбонатной щелочности пока не израсходован Са2+, а растворение СаСО3 удерживало рН> 5.5. Отложение карбонатов в эпоху исключительного господства бактерий происходило в их сообществах вне клеток, что хорошо документировано строматолитами. Бактериальный цикл кальция, оставивший геологические следы в виде карбонатных платформ докембрия, недостаточно понят. Основным последствием его явилось создание нейтральной среды в области развития биосферы (Заварзин, 2002). Геохимический цикл кальция резко изменился с появлением эвкариот, способных к отложению карбонатов внутри клетки под действием карбоанги-дразы, фермента, определяющего скорость установления карбонатного равновесия. Накопление карбонатов стало зависеть от растворения, а не от осаждения.
Субаэральное химическое выветривание шпатов, как основного материала изверженных пород, ведет к образованию карбонатных и содовых условий по общей схеме (рис. 2).
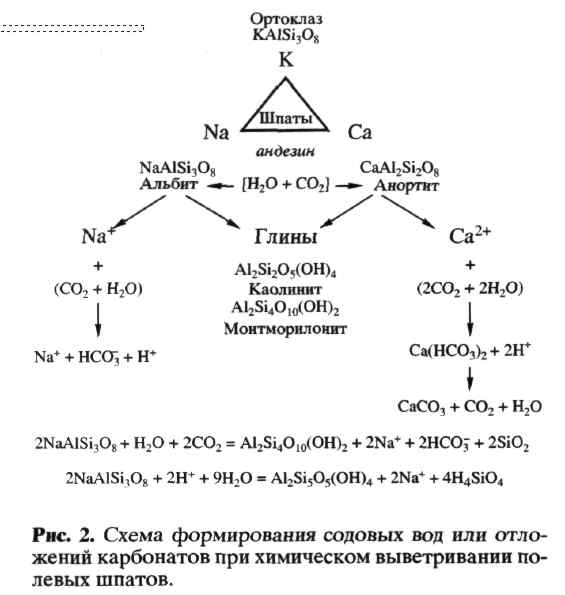
Биотическое влияние на химическое выветривание обусловлено универсальными механизмами метаболического образования Н+ и СО2, как представлено на рис. 2 репрезентативными реакциями. Следствием представленной схемы выветривания являются два пути образования субаэральных условий обитания микробиоты: кальциевый для нейтрофилов и содовый для алкалофилов. Содовые условия могут удерживаться при отсутствии промывного режима в теплых внутрикон-тинентальных областях " содового континента" в семиаридном климате. Отсюда специальный интерес к алкалофильной микробиоте содовых озер как аналогов континентальной биоты в ее противопоставлении морской талассофильной (Заварзин, 1993; Заварзин и др., 1999; Заварзин, Жилина, 2000). Отдельный вопрос представляет ее отношение к эпиконтинентальным морям талассократиче-ских эпох при взаимодействии в области экотона.
Прокариотная биосфера. Главные изменения в геохимии планеты сводятся к формированию системы биогеохимических циклов, катализируемых бактериями. Циклы неполностью замкнуты и ведут к накоплению остаточных продуктов, которые обусловливают биогеохимическую сукцессию как центральное явление, определяющее судьбу биосферы. Конечным результатом служит создание оксической атмосферы с замыканием цикла фотосинтеза дыханием. Деструкция мортмассы ведет к накоплению СО2 в местах деструкции, что ускоряет углекислотное выветривание и, следовательно, связывание СО2 горными породами в маршруте " изверженные породы —-—»• глины + карбонаты". Удаление СО2 из атмосферы снижает парниковый эффект. Образование тонкодисперсных пород приводит к захоронению Сорг, который эквивалентен остаточному О2 атмосферы. Отсюда маршрут " выветривание —»• —»• седиментогенез" с захоронением керогена ответственен за образование кислородной атмосферы и оксических субаэральных условий, а маршрут " выветривание —•• карбонаты" - за создание нейтральной среды. Выветривание идет в субаэральных условиях на платформах, и геологическая летопись сводится к самоуничтожению следов процессов. Центральная роль процессов выветривания - седиментогенеза в преобразовании геосферы приводит к тому, что глубокий океан играет роль преимущественно физического резервуара растворенных веществ, поскольку цикл Сорг в нем замкнут. Отдельно следует рассматривать гидротермальные процессы на дне океана как источник газов и преобразования базальтов, включая их карбонатизацию. Седиментогенез локализован преимущественно в относительно мелководных водоемах. Поэтому необходимо признать гетерогенность географической оболочки Земли с областями интенсивных процессов преобразования на крупных геохимических барьерах и отказаться от попыток распространения данных по одному тафоценозу на всю планету.
Биотические процессы определяют цикл органического углерода. Ключевой реакцией служит образование биомассы при фотосинтезе, пропорциональное освещаемой дневной поверхности. Реакция ассимиляции с самого начала осуществлялась по рибулозобисфосфатному пути, как об этом можно судить по изотопному составу керогена древних пород с обогащением легким изотопом углерода близким 25 х 10~3. Какова была освещенная поверхность, пригодная для цианобак-терий? По-видимому, она не сильно отличалась от суммы площадей воды и суши, занятых фитопланктоном и растениями в настоящее время. Цианобактериальный мат относится к биоценозам организмов с коротким жизненным циклом, быстро набирающим максимальную плотность до полного самозатенения, т.е. около 500-1000 мг хлорофилла/м2. Отсюда можно заключить, что ассимиляция С-СО2 составляла величину п х х 102 млрд. т С/год, где п составляет 1-2. При использовании современных значений для стока С-СО2 в ~ 145 кг/1кг хлорофилла проективного покрытия растительного покрова необходимо сделать поправки на стадии стока углерода соответственно времени пребывания, где стадиям соответствуют:
Gross Primary Production (GPP) - вся ассимилированная углекислота;
Netto Primary Production (NPP) - углерод первичного продуцента за год (фотоассимиляция минус дыхание растения, в том числе ночью, предполагается GPP = 2NPP);
Netto Ecosystem Production (NEP) - экосистем-ная продукция соответствует NPP минус дыхание органотрофов в экосистеме в течение года;
Netto Biome Production (NBP) - накопление углерода в ландшафте в течение десятилетий с образованием устойчивого органического вещества гумуса с временем пребывания ~ 1000 лет.
Для наших целей необходимо ввести понятие нетто-геосферной продукции Netto Geospheric Production (NGP), соответствующее захороненному углероду керогена и каустобиолитов с временем пребывания более 108 лет (геологический рецикл).
В избранном нами масштабе времени все остальные величины, кроме NGP, представляют малые величины динамических резервуаров с наиболее крупной величиной - устойчивым углеродом гумуса, куда входит и растворенное органическое вещество океана. Считается, что для водных экосистем logNGP = klogGPP и количество углерода, переходящего в устойчивую форму в осадках, пропорционально первичной продукции (Tyson, 1995), поскольку для водорослей с коротким жизненным циклом GPP ~ NPP. Из цикла выводится только устойчивый углерод, и таким образом скорость седиментации тонкодисперсных осадков глин контролирует создание кислородной атмосферы (см. рис. 1).
Ключевым этапом для начала образования ке-рогена в осадках фанерозоя служит переход из оксической в аноксическую зону с резким уменьшением скорости деструкции. Эта концепция явно непригодна для ранних этапов эволюции геосферно-биосферной системы, поскольку в атмосфере не было О2. Кислородная обстановка тогда создавалась в экосистемах на короткое дневное время, и в плотных скоплениях водорослей она могла достигать 100% вблизи пузырьков газа. Соответственно бактерии в непосредственной близости от цианобактерий должны были обладать окситолерантностью, хотя бы к стрессовым воздействиям кислорода, а возможно, и быть " half time" аэробами. Круглосуточный аэробиоз вряд ли имел место, а при стоке кислорода в океан создавались условия для микроаэрофилов, каковы многие водные микроорганизмы.
Ассимиляция СО2 происходит в молярном отношении СО2: Сорг: О2 = 1: 1: 1 по реакции СО2 + + Н2О = [СН2О] + О2. Реакция ассимиляции уравновешивается дыханием, осуществляющим обратную реакцию.
С циклом углерода связаны циклы азота и фосфора в отношении, определяемом их включением в биомассу. Для микробов отношение C: N: P = = 106: 16: 1.
В современных условиях лимитирующим биогеном чаще всего служит связанный азот. В про-кариотной атмосфере это было не так, поскольку многие прокариоты способны к ассимиляции N2, особенно в аноксических условиях. Ассимиляция N2 ведет к существенным дополнительным энергетическим затратам, но она не была безусловным ограничением. Таким образом, накопление связанного азота наряду с созданием кислородной атмосферы было необходимым предварительным условием для возможности появления эукариот-протист с фотосинтезом и дыханием как единственными путями метаболизма. Взаимодействие биогеохимических циклов показано на схеме (рис. 3).
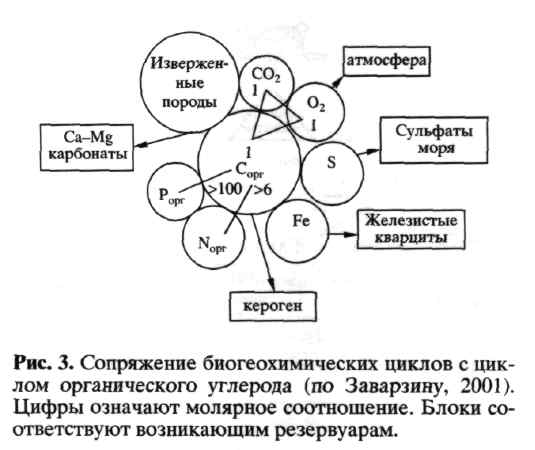
Если цикл Сорг теоретически может быть замкнут растениями (водорослями в океане) и животными, то циклы N, S, Fe катализируются исключительно бактериями. Для цикла азота это цепь: азотфиксаторы - аммонификаторы - нит-рификаторы - денитрификаторы. Для цикла серы: сульфат и сероредукторы (сульфидогены) -серобактерии (анаэробные фототрофные и аэробные тионовые). Для цикла железа: аэробные железобактерии и анаэробные железоредукто-ры. В цикле Сорг в наземных условиях сейчас животные контролируют только 3% от потока углерода, роль основных деструкторов играют почвенные грибы и бактерии; в море положение иное вследствие первичных продуцентов с коротким жизненным циклом и зоотрофными организмами с более длительным временем пребывания Сорг в них по мере прохождения по пищевой пирамиде.
Лимитирующим биогеном для синтеза биомассы был, вероятно, фосфор, который поступает в оборот в результате выветривания и выводится из регенерационного цикла деструкции реакциями с железом и кальцием. В условиях содового континента Fe и Са связаны в нерастворимые соединения, и поэтому концентрация растворенного фосфора может быть выше. Роль цианобакте-риальных сообществ в образовании осадочных месторождений фосфоритов сейчас показана достаточно убедительно (Герасименко и др., 1999).
В цикле органического углерода следует обратить внимание на события, связанные с резким облегчением до -40 и -50 промилль изотопного состава керогена (формации Фортескью, ~2.7 млрд. лет, и Франсвиль, ~2 млрд. лет назад). Эти события интерпретируются как результат накопления биомассы метанотрофов, которые используют метилотрофный путь ассимиляции (Schidlowski, 2002). Если метан может иметь эндогенное происхождение и предположение о метановой атмосфере ранней Земли существует, то для метанотрофов необходим О2, появление которого тогда надо отнести к очень раннему времени.
Микробное сообщество. Для существования в течение геологического времени микробное сообщество должно быть устойчивым. Устойчивость сообщества определяется замыканием трофических связей в цикл таким образом, что конечным продуктом сообщества служит вещество или вещества, поступающие в трофическую цепь в качестве субстратов. Такое сообщество назовем автономным. В автономном сообществе действуют регенерационные циклы для биогенов. Регенерация осуществляется в результате деструкции (Заварзин, Колотилова, 2001). Биологи называют результат деструкции " минерализацией", что не очень удобно для геологической микробиологии, где под минерализацией имеется в виду образование биогенных минералов. Неполная замкнутость циклов ведет к сукцессии. Невозвращенные в регенерационном цикле биогены поступают за счет пополняющих реакций, связанных с выщелачиванием изверженных и осадочных пород.
Трофические связи в прокариотных сообществах были изучены на примере реликтовых сообществ. При их рассмотрении следует иметь в виду, что реликтовые сообщества развиваются в экстремальных условиях, отличных и от современной нормы и от нормы прошлого. Каждое из этих сообществ может служить примером определенных геологических условий. Термофильные сообщества газогидротерм были использованы для моделирования трансформации продуктов вулканических эксгаляций в воздух (Заварзин и др., 1989). Галофильные сообщества, развивающиеся в местах образования эвапоритов, оказались наиболее удобными для исследования путей формирования строматолитов в талассофильных условиях (Заварзин и др., 1993). Новый подход представили сообщества содовых озер (Заварзин и др., 1999). Предполагается, что субаэральное выщелачивание должно вести хотя бы к временному образованию щелочных водоемов, которые таким образом могут служить моделью аталассо-фильных условий. Эти сообщества можно распределить по условному континенту: термофильные сообщества связаны с вулканизмом активного края, галофильные - с областью седиментации на пассивном крае, алкалофильные - относятся к интраконтинентальным областям.
Все цианобактериальные сообщества " матов" построены по одному плану, обусловленному первичными продуцентами - цианобактериями. Они создают плотный слой на поверхности, который при толщине 1-2 мм полностью поглощает фотосинтетически активную радиацию. Под этим слоем располагается слой деструкции в 4-5 мм. В трофической цепи анаэробной деструкции преимущественное значение имеет продукт-субстратное взаимодействие, когда продукт одной группы организмов служит субстратом последующей. Трофическое взаимодействие между микроорганизмами определяется правилом минимального диффузионного расстояния. Наиболее выгодным оказывается построение сообщества из тонких слоев последовательно вступающих в дело группировок организмов. В результате получается тонкослоистая система в 1-4 мм. Поэтому парагистология матов, или их архитектура, универсальна, хотя виды организмов, занимающие каждую из функциональных ячеек в трофической системе, различны и приспособлены к своим топическим условиям.
Трофическая система мата также имеет универсальную структуру с последовательностью определенных групп организмов, начиная с гид-ролитиков, разлагающих частицы мортмассы, и кончая вторичными анаэробами. Важным свойством этой системы оказался тот факт, что наиболее тесно трофически взаимодействующие группы организмов относятся к очень удаленным ветвям филогенетического дерева. Так, клостри-дии-гидролитики взаимодействуют с диссипотро-фами-спирохетами. Те в свою очередь взаимодействуют с протеобактериями-сульфатредуктора-ми или же с архебактериями-метаногенами. Отсюда следует вывод, что сообщество сформировалось не путем дивергенции предковых форм, а было собрано из разнородных организмов, обладающих необходимыми функциями. Завербованные организмы не обязательно оптимальны, но обязательно совместимы с уже существующим сообществом. Совместимость с прошлым составляет принципиальную особенность аддитивной эволюции.
При рассмотрении трофической системы автономного сообщества важно, чтобы в результате деструкции получались вещества, служащие субстратами первичных продуцентов. Для аэробных организмов система представляет веер трофических маршрутов, считая, что для каждого компонента мортмассы есть специализированный деструктор, окисляющий белки, жиры, углеводы до углекислоты. Для анаэробного сообщества это не так. Здесь трофическая цепь работает как каскад реакций: гидролитики - первичные анаэробы - вторичные анаэробы. Конечными продуктами здесь оказываются либо метан в пресных условиях, либо сероводород в минерализованных водах. Соответственно необходимо установить пути регенерации этих продуктов в субстраты. В современной оксической атмосфере метан окисляется метанотрофами, сероводород -тионовыми и серобактериями. Альтернативным путем для метана служит его фотохимическое окисление, имеющее в современной атмосфере мощность 500 Мт в год. Как оно происходило в аноксической атмосфере - неясно. Сероводород может удаляться, связываясь с железом в сульфиды. Для ранней биосферы есть геологические свидетельства маршрутов от Fe(II) к Fe2O3 + Fe3O4 и от H2S к H2SO4, относящиеся к образованию железорудных формаций и сульфатов океана, причем каждый из них послужил за историю Земли стоком для 42-45% О2, рассчитанного как эквивалент керогена осадочных отложений.
При рассмотрении трофической системы в аноксических условиях возможным путем замыкания циклов является деятельность аноксигенных фототрофов - пурпурных и зеленых бактерий. При этом так называемые " несерные фототрофы" способны использовать несбраживаемые продукты первичных брожений, а " серные фототрофы" - окислять сероводород в сульфаты. Таким образом, " пурпурные бактерии", относимые к началу филогенетического дерева, оказываются в конце трофической цепи, завершая процесс деструкции. Загадкой до последнего времени оставалось анаэробное окисление метана. Фототрофы к анаэробному окислению метана неспособны. Специалисты из Института морской микробиологии (Бремен) установили, что анаэробное окисление метана происходит синтрофно в консорциуме с участием метаногенов и сульфатредукторов. Им же принадлежит открытие фототрофного анаэробного окисления железа чистой культурой несерных пурпурных бактерий. Отсюда возникает потенциальная возможность замыкания циклов железа и серы в аноксических условиях. Общую картину трофических взаимодействий в бактериальном сообществе с участием фототрофов можно представить сокращенной схемой (рис. 4).
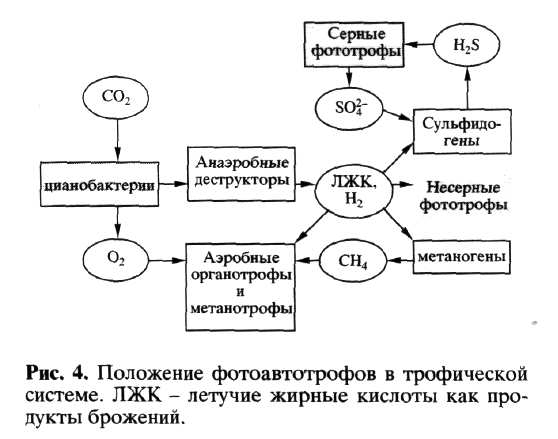
Блок бактериальной деструкции на рис. 4 представлен в обобщенном виде, на самом деле он представляет кооперативное сообщество со сложными трофическими связями (Заварзин, Ко-лотилова, 2001). Из схемы трофических отношений следует, что в современных сообществах аноксигенные фототрофы, которых филогенетики ставят в начало эволюционной линии, на самом деле замыкают путь деструкции с каталитической ролью серного цикла в анаэробных условиях. Возможно ли обращение этой схемы справа налево с аноксигенными фототрофами как первичными продуцентами, если Н2 и H2S поступают извне из абиогенных источников? Для современных сообществ такие примеры неизвестны.
Становление биогеохимических циклов. Для становления биогеохимических циклов необходимо участие разнообразных групп прокариот, взаимодействующих друг с другом. Реакции геохимических циклов сопряжены друг с другом в единую систему. Прокариоты осуществляют катализ важных геохимических реакций, при относительно малом вкладе собственной биомассы. Их концентрационная функция в отличие от скелетных организмов мала и связана с биологически опосредованными реакциями, когда деятельность микроорганизмов приводит к созданию условий, в которых образуются минералы, такие как карбонаты, сульфиды, окислы железа, фосфаты, отложения которых установлены в ранние периоды истории Земли. Противопоставление преимущественно каталитической активности микробной системы и концентрационной деятельности биоты, на которую ориентировался Вернадский, существенно для прокариотной биосферы. Циано-бактерии, казалось бы способные развиваться независимо от других организмов, имеют два существенных ограничения. Во-первых, они неспособны развиваться в условиях повышенного содержания О2 выше 20%, что служит для них пределом самоотравления продуктами. Лучше они растут в аноксических или даже восстановительных условиях. Во-вторых, они неспособны осуществлять регенерационный цикл, для которого необходима описанная выше кооперативная система микроорганизмов.
Из необходимости циклов Вернадский сделал вывод, к которому полностью должен присоединиться микробиолог, занимающийся функциональным разнообразием: невозможно построить устойчивую систему на монофилетическом основании. Универсальный предок не может существовать, поскольку он представлял бы собой " вечный двигатель", даже будучи фототрофом - автотроф-ным днем и гетеротрофным ночью. Представление об " универсальных предках" логически противоречиво. Изначальная группировка микроорганизмов должна была быть разнородной и функционально комплементарной. Иные группировки не могли быть устойчивыми и существовать в течение геологически длительного времени. Этим требованиям отвечает цианобактериальное сообщество. Бактериальная палеонтология в общем, со всеми ссылками на неполноту геологической летописи, на трудность идентификации функции по микрофоссилиям, подтверждает вывод о древности цианобактериального сообщества (Сергеев и др., 2002). Топология филогенетических бескорневых деревьев указывает на циано-бактериальную группировку как на " сердцевину подсолнечника", из которой радиусами расходятся линии (Zavarzin et al., 1990). Но сила этого аргумента ослабляется тем обстоятельством, что мо-лекулярно-биологическая классификация построена на основании ныне живущих бактерий, которые совместимы с оксигенными первичными продуцентами.
Существенно, что прокариотная система самодостаточна и для поддержания собственного существования в течение неопределенно долгого времени (фактически - в течение всей геологической истории Земли), и для осуществления биогеохимической сукцессии, составляющей историю биосферы и суть ее эволюции. Усложнение организмов в их филогенетическом пути происходит в условиях сложившейся и стабилизированной про-кариотной биосферы с модификацией некоторых реакций за счет морфологического усложнения, в первую очередь транспорта. Можно рассматривать три этапа эволюции: обмен небольшими участками мозаичного генома у прокариот, системную комбинаторику эндосимбиоза у протист, генеалогическую картину последовательного усложнения близкого к монофилетическому дарвинистскому дереву сложных организмов (Завар-зин, 2002). Переходу к эвкариотным протистам с фаготрофным питанием должно было предшествовать появление цитологически парадоксального " фаготрофного прокариота" (Cavalier-Smith, 2002), и лишь за этим последовали симбиогенети-ческие события.
Применение актуалистического подхода к прошлому предполагает персистентность организмов и служит постоянным источником возражений со стороны генетиков, утверждающих, что нельзя говорить об идентичности морфологически сходных организмов, особенно прокариот, например трихомных цианобактерий и серобактерий. Однако противоположное утверждение о различии между древними и нынешними организмами автоматически отправляет в корзину все филогенетические деревья, построенные на основе гомологии последовательностей участков генома современных организмов. Вряд ли это вполне разумно.
Коротко нужно отвлечься на описание выпадающих из общей картины эволюции прокариотной биосферы глубинных гидрогенотрофных хемосинтезирующих организмов. Источником питания для них служат дающие водород реакции воды с перегретыми горными породами. Проблему составляют акцепторы электрона, которыми могут служить углекислота или соединения серы. Продуктами обмена глубинных микроорганизмов являются соответственно метан или сероводород. Выгодная область развития этих организмов, связанных с перегретыми породами, - зона конденсации воды над магматическими очагами. Поэтому многие из них относятся к гипертермофилам. Группировка хемосинтезирующих микроорганизмов, не зависящих от солнечного света, безусловно принадлежит к древнейшей и реликтовой в полном смысле слова. Но гидрогенотрофные организмы находятся в стороне от магистрального пути эволюции биоты. Их важной функцией могло быть связывание водорода, который не мог диссипировать в космос и приводить к окислительной гибели планеты, как это произошло на Марсе и Венере. Такая обратная связь от микробиоты к геосфере пока не была рассмотрена.
Следует заметить, что термофильных окси-генных фототрофов нет, верхний предел их развития лежит между 60-70°С, в то время как нижний предел лежит около 0°С, что существенно для цикла органического углерода.
На поверхности Земли изначально для геологической истории действовало микробное сообщество, сопрягающее циклы СО2, 02, Сорг, N, P, S, Fe в реакциях, связанных с синтезом биомассы и катаболизмом микроорганизмов, а также осуществляющее преобразование алюмосиликатных пород, кальция, магния, кремния - в биологически опосредованных реакциях. Циклы азота, серы, железа, в которых участвуют специфические функциональные группировки прокариотных микроорганизмов, здесь не разбирались, точно так же как реакции фосфора. Они достаточно хорошо известны в природоведческой микробиологии.
Прокариотная биота эволюционировала на основе комбинаторного перераспределения экологически значимых функций между филогенетически различными организмами, условием выживания для которых было занятие функциональной ниши в существующей системе отношений в сообществе. Морфологические приспособления при этом также имели значение: трихомные организмы приурочены к илу, мицелиальные - к суб-аэральной среде. Геосферно-биосферная система в целом эволюционировала за счет биогеохимической сукцесии как главной движущей силы, ведущей к изменению среды обитания. Наиболее устойчивым временем существования был мезо-протерозой, с окончанием которого и становлением биосферных циклов современного типа сложились предпосылки для субститутивной эволюции с ключевой ролью первичных продуцентов -водорослей, а затем и наземных растений. Дальнейшая эволюция биоты путем интегративного симбиогенеза и последующего усложнения вела к сужению базы жизни для высших форм и ограничению ее " нормальными" условиями. Поддержание всей геосферно-биосферной системы было возможно лишь за счет сохранения системы трофических отношений прокариот как основы биосферы. Появление фаготрофного питания протист, а затем " Parazoa", укоротило пути деградации взвешенного органического вещества. При этом большую роль играли симбиотические отношения хозяев с инкорпорированным микробным сообществом. У зоотрофных животных, способных питаться живыми фототрофными продуцентами, они достигли максимума с образованием микробного сообщества в пищеварительном тракте.
В результате открытий последних десятилетий, которые можно отнести к бактериальной палеонтологии в широком смысле слова, представления об эволюции геосферно-биосферной системы дали основу новому мировоззрению, в котором большее значение придается кооперативным взаимоотношениям в рамках большой системы. В этом отношении рассмотренная система находится вне области традиционных для биологов популяционных пределов, составляющих основу дарвинизма. Одномоментность как необходимое условие взаимодействия возвращает рассмотрение к географической системе А. Гумбольдта. Поэтому рассматриваемую область удобно обозначить как внедарвинистскую область геосфер-но-биосферных систем. В ней наряду с прямыми связями от геосферы к биоте большую роль играют трансформирующие обратные связи в большой системе.
СПИСОК ЛИТЕРАТУРЫ
Виноградский С.Н. Круговорот жизни (1896) // Вести. РАН. 1996. Т. 66. № 12. С. 1115-1120.
Герасименко Л.М., Заварзин Г.А., Розанов А.Ю., Ушатинская Г.Т. Роль цианобактерий в образовании фосфатных минералов // Журн. общ. биол. 1999. Т. 60. № 4. С. 415-430.
Заварзин Г.А. Бактерии и состав атмосферы. М.: Наука, 1984. 193 с.
Заварзин Г.А. Эпиконтинентальные содовые водоемы как предполагаемые реликтовые битопы формирования наземной биоты // Микробиол. 1993. Т. 62. № 5. С. 789-800.
Заварзин Г.А. Становление биосферы // Вести. РАН. 2001. Т. 71. № 11. С. 988-1001.
Заварзин Г.А. Роль комбинаторных событий в развитии биоразнообразия // Природа. 2002. № 1. С. 12-19.
Заварзин Г.А., Герасименко Л.М., Жилина Т.Н. Циано-бактериальные сообщества в гиперсоленых лагунах Сиваша // Микробиол. 1993. Т. 62. № 6. С. 579-599.
Заварзин Г.А., Жилина Т.Н. Содовые озера как природная модель древней биосферы // Природа. 2000. № 2. С. 44-55.
Заварзин Г.А., Жилина Т.Н., Кевбрин В.В. Алкало-фильное микробное сообщество и его функциональное разнообразие // Микробиол. 1999. Т. 68. № 5. С. 503-521.
Заварзин Г.А., Карпов Г.А., Горленко В.М. и др. Каль-дерные микроорганизмы. М.: Наука, 1989. 120 с.
Заварзин Г.А., Колотилова Н.Н. Введение в природоведческую микробиологию. М.: МГУ, 2001.
Сергеев В.Н., Герасименко Л.М., Заварзин ГА. Протерозойская история цианобактерий и их современное состояние // Микробиол. 2002. Т. 71. № 6. С. 725-740.
Сорохтин О.Г., Ушаков С.Д. Развитие Земли. М.: МГУ, 2002.
Хаин В.Е., Короновский Н.В., Ясаманов Н.А. Историческая геология. М.: МГУ, 1997.
Cavalier-Smith T. The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa // Int. J. Syst. Evol. Microbiol. 2002. V. 52. P. 297-354.
Franch S., Bounama C. Effects of water-dependent creep rate on the volatile exchange between mantle and surface reservoirs // Phys. Earth and planetary interiors. 1995. V. 92. P. 57-65.
Lovelock J.E. The ages of Gaia - a biography of living Earth (2d ed.). Oxford: Oxford Univ. Press, 1995.
Schidlowski M. Sedimentary carbon isotope archives as recorders of early life: implications for extraterrestrial scenarios // Fundamentals of life / Eds. G. Palyi, C. Zucchi, L. Ca-glioti. Amsterdam: Elsevier, 2002. P. 307-329.
Schwartzman D.W. Life, temperature and the Earth: the self-organizing biosphere. N. Y.: Columbia Univ. Press, 1999.
Tyson R.V. Sedimentary organic matter. Organic facies and palinofacies. L.: Chapman and Hall, 1995. 615 p.
Westall F., de Wit MJ., Dann J. et al. Early Archean fossil bacteria and biofilms in hydromemally-influenced sediments from the Barberton greenstone belt, South Africa // Precam-brian Res. 2001. V. 106. P. 93-116.
Zavarzin GA., Stackebrandt E., Murray R.G.E. A correlation of phylogenegenetic diversity in the Proteobacteria with the influences of ecological forces // Canad. J. Microbiol. 1990. V. 37. P. 1-6.