Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Електричний синапс
|
|
Будова
Переважна більшість синапсів в нервовій системі царства тварин є саме хімічними. Для них характерна наявність кількох спільних рис, хоча, тим не менше, розміри та форма пре- та постсинаптичних компонентів варіюють дуже широко.
Пресинаптичний пухирець містить численні синаптичні везикули, в яких знаходиться медіатор. Їхній однаковий розмір у всіх досліджених синапсах (40—50 нанометрів) спочатку вважали доказом того, що кожна везикула є мінімальним кластером, чиє вивільнення потрібне для продукування синаптичного сигналу. Везикули розміщуються навпроти пресинаптичної мембрани, що обумовлено їхнім функціональним призначенням для вивільнення медіатора в синаптичну щілину. Також в пресинаптичному пухирці наявна велика кількість мітохондрій (що виробляють аденозинтрифосфат) та впорядковані структури протеїнових волокон.
Синаптична щілина — це простір між пресинаптичним пухирцем та постсинаптичною мембраною від 20 до 30 нанометрів завширшки, що містить зв'язуючі пре- та постсинапс структури, побудовані з протеоглікану. Ширина синаптичної щілини в кожному окремому випадку обумовлена тим, що вивільнений з пресинапсу медіатор має проходити до постсинапсу за час, що є впевнено меншим за частоту нервових сигналів, характерних для нейронів, що утворюють синапс (час проходження медіатору від пре- до постсинаптичної мембрани — звичайно близько кількох мікросекунд).
Постсинаптична мембрана належить клітині, що приймає нервові імпульси. Механізмом трансляції хімічного сигналу медіатора в електричний потенціал дії на цій клітині є рецептори — білкові макромолекули, що вбудовані в постсинаптичну мембрану.
За допомогою спеціальних ультрамікроскопічних методик в останні роки було отримано досить великий обсяг інформації щодо детальної структури синапсів.
Так, було відкрито впорядковані струтури кратероподібних заглиблень діаметром 10 нанометрів, що є вдавлені всередину та перфорують пресинаптичну мембрану. Спочатку їх було названо синаптопорами, але зараз ці структури називають місцями приєднання везикул (МПВ). МПВ зібрані у впорядковані групи чисельністю по шість окремих поглиблень навколо так званих ущільнених виступів (див. схему та мікрофотографію). Таким чином, ущільнені виступи формують правильні трикутні структури на внутрішній стороні пресинаптичної мембрани, а МПВ — гексагональні, і є місцями, де везикули відкриваються і викидають медіатор у синаптичну щілину.
2.2. Механізм передачі нервового імпульсу
Надходження електричного імпульсу до пресинаптичного пухирця вмикає процес синаптичної передачі, першим етапом якої є входження іонів Са2+ ззовні до пресинапсу через спеціалізовані кальцієві канали, що локалізовані біля синаптичної щілини. Са2+ за допомогою невідомого поки що повністю механізму активує везикули, які сукупчені біля МПВ, а ті вивільняють медіатор в синаптичну щілину. Іони Са2+, після активації ними везикул з медіатором, миттєво (за час порядку кількох мікросекунд) деактивуються завдяки депонуванню в мітохондріях та везикулах пресинапсу.
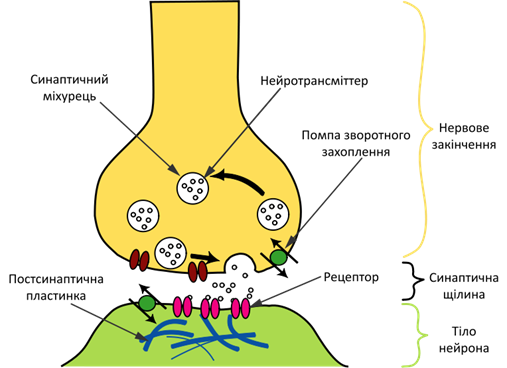
Рис.1 Схема процесу передачі нервового сигналу в хімічному синапсі
Молекули медіатора, що вивільняються з пресинапсу, зв'язуються з рецепторами на постсинаптичній мембрані, в результаті чого в рецепторних макромолекулах відкриваються іонні канали (у випадку канальних рецепторів, що є найбільш розповсюдженим їхнім типом; при роботі рецепторів інших типів механізм передачі сигналу відрізняється). Іони, що починають надходити всередину постсинаптичної клітини через відкриті канали, змінюють заряд її мембрани, що призводить до часткової деполярізації мембрани і, як наслідок, провокування генерації постсинаптичною клітиною потенціалу дії (у випадку впливу на постсинаптичну клітину збуджувальних синапсів), або гіперполярізації мембрани і, таким чином, блокування генерації потенціалу дії (у випадку впливу гальмівних синапсів).
2.2.1 Квантово-везикулярна гіпотеза
Найбільш поширена до останнього часу в якості пояснення механізму вивільнення медіатора з пресинапса гіпотеза квантово-везикулярного екзоцитозу (КВЕ) має на увазі, що «пакет», або квант медіатору міститься в одній везикулі і вивільняється при екзоцитозі (при цьому мембрана везикули зливається з клітинною пресинаптичною мембраною). Це довгий час було превалюючою гіпотезою — незважаючи на той факт, що кореляція між рівнем вивільнення медіатора (або постсинаптичними потенціалами) та кількістю везикул в пресинапсі відсутня. Окрім того, гіпотеза КВЕ має і інші істотні недоліки.
Фізіологічним підґрунтям саме квантованого вивільнення медіатора має бути однакова кількість цього медіатора в кожній везикулі. Гіпотеза КВЕ в класичному вигляді не пристосована до опису ефектів квантів різного розміру (або різної кількості медіатора) що можуть бути вивільнені при одному акті екзоцитозу. При цьому треба прийняти до уваги, що в одному і тому ж пресинаптичному бутоні можуть спостерігатись везикули різного розміру; окрім того, не знайдено кореляції між розміром везикули та кількістю медіатора в ній (тобто його концентрація в везикулах теж може бути різною). Більш того, в денервованому нервово-м'язевому з'єднанні (або нервово-м'язевому синапсі) шванівські клітини генерують більшу кількість мініатюрних постсинаптичних потенціалів, аніж спостерігається в синапсі до денервації, незважаючи на повну відсутність пресинаптичних везикул в цих клітинах, локалізованих в районі пресинаптичного бутону.
2.2.2. Гіпотеза пороцитозу
Існують істотні експериментальні підтвердження про те, що медіатор секретується в синаптичну щілину завдяки синхронній активації гексагональних груп МПВ (див. вище) та приєднаних до них везикул [3], що стало підґрунтям для формулювання гіпотези пороцитозу. Ця гіпотеза базується на спостереженні, що приєднані до МПВ везикули при надходженні потенціалу дії синхронно скорочуються і при цьому секретують в синаптичну щілину щоразу однакову кількість медіатора, вивільняючи тільки частину вмісту кожної з шести везикул. Сам по собі термін «пороцитоз» походить від грецьких слів poro (що означає пори) та cytosis (описує перенос хімічних субстанцій через плазматичну мембрану клітини).
Більшість експериментальних даних щодо функціонування моносинаптичних міжклітинних з'єднань отримано завдяки дослідженням ізольованих нервово-м'язевих контактів. Як і в міжнейронних, в нервово-м'язевих синапсах МПВ формують впорядковані гексагональні структури. Кожна з таких гексагональних структур може бути визначена як «синаптомер» — тобто структура, яка є елементарною одиницею в процесі секреції медіатора. Синаптомер містить, окрім власне порових заглиблень, протеїнові нитчаті структури, які утримують везикули лінійно впорядкованими; існування аналогічних структур доведено і для синапсів в центральній нервовій системі (ЦНС).
2.3. Види хімічних синапсів
На сучасному етапі вивчення синапсів все ще не стало можливим вироблення загальної класифікації цих структур на структурно-функціональному підґрунті. Анатомічно синапси можуть знаходитись на дендритних стовбурах, на кінцевих дендритних відгалуженнях, на тілі нейрону, або навіть на аксональних терміналах інших синапсів — в такому випадку до процесу передачі сигналу залучаються рецептори і на пресинаптичній мембрані іншого синапсу. Чітка локалізація гальмівних та збуджувальних синапсів на різних частинах нейрону простежується лише в деяких випадках. Наприклад, в спинному мозку пірамідальні нейрони утворюють інгібіторні (гальмівні) синапси на тілі клітини, а збуджувальні — на кінцевих відгалудженнях дендритів. Аналогічне анатомічне впорядкування характерне для клітин Пуркін'є в мозочку. Але, загалом, всеоохоплюючих закономірностей розташування синапсів на нейроні наразі не сформульовано.
Завдяки забарвленню іонами важких металів синапси були поділені на два типи, що часто називаються Gray-1 та Gray-2. Для типу 1 характерна наявність інтенсивного постсинаптичного ущільнення і синаптична щілина шириною 30 нанометрів; ущільнення розповсюджене практично на всю постсинаптичну мембрану (див. мікрофотографію). Тип 2 демонструє набагато менш виражене забарвлене ущільнення, і ширину синаптичної щілини, що дорівнює 20 нанометрам. Як показано в дослідах на гіпокампі та головному мозку, локалізовані там синапси типу 1 є збуджувальними, а типу 2 — гальмівними. За деякими даними, ця кореляція спостерігається також в корі та в інших частинах головного мозку ссавців, але в спинному мозку майже не спостерігається.
Надалі було показано, що початкова альдегідна фіксація препаратів мозку надає (з невідомих досі причин) синаптичним везикулам сплюснутої форми, але тільки в синапсах типу 2; радіолігандне мічення показало, що в таких синапсах із сплюснутими везикулами в якості медіатора (а, отже, вмісту везикул) використовується гамма-аміномасляна кислота (ГАМК) або гліцин — тобто інгібіторні (гальмівні) нейромедіатори. Таким чином було встановлено додатковий структурно-функціональний зв'язок для частини синапсів.
Окрім того, синапси можуть бути класифікованини за медіатором, який вони використовують ГАМК, ацетилхолін, норадреналін, опіати і т. ін.
2.4. Головні особливості хімічних синапсів
1. Синтез нейромедіатора у пресинаптичному нервовому закінченні.
2. Накопичення нейромедіатора в секреторних пухирцях.
3. Регульоване вивільнення нейромедіатора в синаптичну щілину.
4. Наявність специфічних рецепторів для нейромедіатора на постсинаптичній мембрані, які зумовлюють відповідний ефект при дії на них нейромедіатора. Аплікація нейромедіатора на синапс імітує наслідки подразнення нерва.
5. Наявність механізмів, що припиняють дію вивільненого нейромедіатора.
Електричний синапс
|
|