Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Лекция жоспары. 1. Потенциал туралы түсінік
|
|
1. Потенциал туралы тү сінік
2. Биопотенциал тү рлері: тыныштық жә не ә сер потенциалдары
3. Гольдман-Ходжкин-Катц тең деуі.
4. Ә рекет потенциалының пайда болу механизмі.
5. Ходжкин-Хаксли тең деуі.
6. Ә рекет потенциалының таралуы.
Лекция мақ саты: биопотенциалдың тү рлерін, пайда болу механизімін, оның математикалық ө рнегін жә не ә рекет потенциалының нерв талшығ ымен таралуын талдау.
Физикада потенциал термині ә р тү рлі ө рістердің (электростатикалық, магниттік, гравитатциялық т.б.) энергиясын сипаттау ү шін қ олданылатын шама. Биологиялық мембранада электр ө рісі бар, олай болса ол ө рістің энергиясын да осы тә різді бипотенциал деген шамамен сипаттаймыз. Кө біне потенциалдың шамасын қ олданбайды, олардың айырымын, яғ ни екі нү ктедегі потенциалдар айрымын қ олданады: Dj =j2 - j1, мұ ндағ ы j2 и j1 сә йкес нү ктелердегі (аймақ тардағ ы) потенциал шамасы. Биопотенциал деп ағ заның екі нү ке арасындағ ы потенциалдар айырымын немесе мембранадағ ы биопотенциал деп, мембрананың ішкі жә не сыртқ ы орта арасындағ ы потенциалдар айырымын атайды: DjМ =jІШ - jСЫРТ.
Барлық уақ ытта қ озбағ ан жасушада потенциал айырымы кездеседі, оны тыныштық потенциалы деп атайды, жасуша ішінде потенциал «теріс», сыртында «оң» таң балы болады.
Биопотенциалдың пайда болу механизімін толық талдау ү шін, біз алдымен олардың негізінде қ андай физика-химиялық қ ұ былыстардың жатқ андығ ын, жә не потенциалдың шамасы неге байланысты болатындығ ын талдайық.
Пайда болу механизміне байланысты потенциалдар: диффузиялық, фазалық жә не мембраналық деген тү рлерге бө лінеді. Диффузиялық потенциал сұ йық ортаны бө ліп тұ рғ ан шекарадағ ы иондардың қ озғ алғ ыштығ ының ә р болуынан пайда болады (1 сурет).
HCl Н2О  j- j+
j- j+
Н+
Cl-
1 cурет
Мысалы, ортасында поры бар бө гет арқ ылы екіге бө лінген ыдыстың сол жағ ына тұ з қ ышқ ылы ертіндісін (HCl), оң жағ ына су қ ұ яйық. Пор тек иондарды ө ткізсін делік. Диффузия қ ұ былысы ә серінен жә не екі ортада концентрациялық градиенттің болуы себебінен иондар ыдыстың екінші жағ ына қ арай қ озғ алады. Концентрациялық градиент деп белгілі бір бағ ыттағ ы бірлік ұ зындық қ а сә йкес келетін концентрация айрымына тең шаманы атайды. Мысалы, ертіндінің бірінші нү ктедегі концентрациясы С1, одан l қ ашық тық та жатқ ан екінші нү ктедегі концентрациясы С2 болса, онда gradC=С1- С2/ l тең болады.
Сонымен концентрациялық градиенттің нолден ө згеше болуы жә не оң зарядталғ ан сутегі иондарының қ озғ алғ ыштығ ының жоғ ары болуы себепті, оң зарядталғ ан сутегі иондары теріс зарядталғ ан хлор иондарын артқ а тастап, ыдыстың оң жағ ына бұ рын жетеді, нә тижесінде ыдыстың оң жақ бө лігі оң зарядталады, ал сол жақ бө лігінде оң зарядтардың аз болуы (сутегі иондарының біраз бө лігі оң жақ қ а ауысып кетті) теріс зарядтардың басым болуы себепті ол орта теріс зарядталады. Осылайша ыдыстың екі жағ ындағ ы екі тү рлі зарядтардың арасында электр ө рісінің айырымы туындайды оны диффузиялық потенциал деп атаймыз жә не оның шамасы Гендерсон тең деуімен анық талынады:
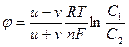
мұ ндағ ы u- катионның жылдамдағ ы, v- анионның жылдамдығ ы, R- газ тұ рақ тысы, T- абсолюттік температура, n- ионның валенттілігі, F- Фарадей саны, c1 – бірінші ортадағ ы ионның концентрациясы, c2 – екініш ортадағ ы ионның концентрациясы. Бұ л ө рнектен, дифффузиялық потенциалдың шамасы катион мен анионның қ озғ алғ ыштығ ының айрымашылығ ына, олардың концентрациясына тікелей байланысты екендігі кө рінеді. Егер катион мен анионның жылдамдығ ы тең болса, немесе олардың концентрациясы бірде болса, онда диффузиялық потенциал нолге тең болады.
Биологиялық жү йелерде потенциалдың бү л тү рі негізінен жасушаның механикалық зақ ымдалуы кезінде байқ алады жә не иондардың зақ ымдалғ ан аймақ тан зақ ымдалмағ ан аймақ қ а қ арай диффузиясы жү реді.
Қ арастырылғ ан мысалдағ ы бө гет аниондарды тіптен ө ткізбесін делік, сонда ыдыстың оң жағ ына тек сутегі иондары ө теді, яғ ни v =0 болғ андық тан
Гендерсон тең деуі Нернст тең деуіне айналады: j= RT/nF ln C1/C2.
Фазалық потенциалдар араласпайтын екі фазалық кү йдегі заттар шекарасында (судағ ы электролит ертіндісі мен май) пайда болады. Мысалы, анионғ а салыстырғ анда катиондар майлы ортада жақ сы еритін болса, онда ол майлы ортадағ а жылдам ауысып, ортаны оң зарядтайды.
Мембраналық потециалдың пайда болуы туралы алғ ашқ ы теорияны 1902 жылы Бернштейн ұ сынды. ХХ ғ асырдың 50 жылдары Ходжикин, Хакси жә не Катц, Бернштейнннің теориясының дұ рыстығ ын тә жірибе арқ ылы дә лелдеді. Олар мембрананың иондарды таң дап ө ткізуінен мембрананың іші мен сыртқ ы орталарындағ ы иондардың концентрациясының ә р тү рлі болуы биопотенциалдың пайда болуының басты себебі деген қ орытынды жасады жә не оны кальмардың аксонында жү ргізген тә жрибелері арқ ылы толық дә леледі. Тә жірибе қ озбағ ан мембрананың (кальмар аксоны) калий иондарын жақ сы, ал натрии жә не басқ а иондарды нашар ө ткізетіндігін кө рсетті. Оның ү стіне калии иондарының концентрациясы жасуша ішінде сыртқ ы ортамен салыстырғ анда 20-40 есе кө п, олай болса, диффузия ә серінен калии иондары сыртқ а қ арай тасымалданады, ал сыртқ ы ортадағ ы иондар ішке ене алмайды, олар ү шін мембрана ө тімділігі ө те тө мен. Сонымен сыртқ ы ортағ а ө ткен калии иондары ортаны оң зарядтайды, ішкі ортадағ а оң зарядтардың біраз бө лігін калий алып кеткендіктен, ішкі орта теріс зарядты болады (ішкі ортадағ ы теріс зарядтар сыртқ а ө те алмайды, оң зарядтар азайды). Бұ л ү рдіс динамикалық тепе тең дік орнағ анғ а дейін жү реді. Сонымен тыныштық потенциалы калий иондарының циоплазмадан сыртқ ы ортағ а қ арай бағ ытталғ ан диффузия ә серінен деп қ арастырсақ, онда оның шамасын Нернст
тең деуінен анық тауғ а болады:
j= RT/nF ln C1/C2
мұ ндағ ы К1 жасушаның ішіндегі, К2 сыртындағ ы калии ионының концентрациясы. Бұ л тең деуді тепе тең дік потенциалы деп те атайды. Кальмар аксонындағ ы тыныштық потенциалының шамасы jМ = - 60мВ (2 сурет).
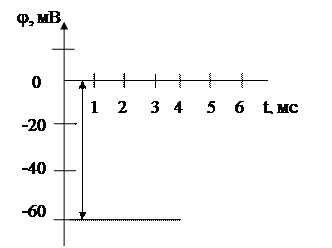 |
jМ = - 60мВ
2 сурет
Кейінгі жү ргізілген тә жірибелер мембрананың аз мө лшерде натрия мен хлор иондарын да ө ткізетіндігін кө рсетті, олай болса тыныштық потенциалының пайда болуында K+, Na+ жә не Cl- иондарынының да қ атысы бар. Қ озбағ ан мембрананың K+, Na+ жә не Cl- иондары ө ткізу коэффиценті PK: PNa: PCl = 1: 0, 04: 0, 45 мұ нан калий ионының натриғ а салыстырғ анда ө тімділігі 25 есе жоғ ары екендігі кө рінеді, яғ ни тыныштық потенциалының пайда болуында калии мен натрии иондарының концентрациялары басты роль атқ аратындығ ы келіп шығ ады.
1949 жылы Гольдман, Ходжкин жә не Катц тыныштық потенциалының басты тең деуін ұ сынды:
jM = -RT/F ln 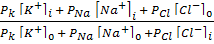 ,
,
мұ ндағ ы i и 0 иондардың ішкі жә не сыртқ ы ортадағ ы концентрациясы, РК, РNa РCl сә йкес иондар ү шін мембрананың ө ткізгіштік коэффиценті. Бұ л ө рнек Гольдман- Ходжкин - Катц тең деуі деп аталады.
Жасушаны сыртқ ы тітіркендіргіштер (механикалық, жылулық, электрлік) арқ ылы қ озғ ан кү йге кө шірсек, мембрананың натрии иондарын ө ткізуі кү рт жоғ арылайды, бірақ калии иондарын ө ткізу дә режесі ө згеріссіз қ алады. Мембрананың калии жә не натрии иондарын ө ткізу коэффициенті PK : PNa = 1: 20 дейін ө згеріп, натрии ионын ө ткізу 500 есе артады. Оның ү стіне мембранада натрии иондарының каналы 10 есе кө п, осы факторлар натрии иондарының мембрана ішіне қ арай ө туін кү шейтіп, ондағ ы потенциалдар айрымын тө мендеуіне, соң ынан оның ішкі қ абатының оң зарядталуына алып келеді. Жасушадағ ы бұ л қ ұ былысты деполяризация деп атайды. Мембрананың қ озу нә тижесіндегі потенциалдар айрымының толық ө згерісі ә рекет потенциалының мә нін береді: jӘ = jТ - Dj, где jТ – тыныштық потенциаланың мә ні, Dj- потенциал ө згірісі. Ә рекет потенциалы деп қ озғ ан кү йдегі цитоплазма мен сыртқ ы орта арасындағ ы потенциалдар айрымын айтамыз. Мысалы, ү лкен кальмар аксонының тыныштық потенциалы -60 Мв тең. Оны қ оздырғ анда потенцилы + 35 мВ дейін жоғ арыласа, онда потенцилдың айырымы немесе ә рекет потенциалы jӘ = - 60 мВ – 35 мВ = - 95 мВ (3- сурет).
 |
Dj
1 2 3 4 5 6 t, мс
jT jӘ
t
3- сурет. jТ – тыныштық потенциалы, jӘ – ә рекет потенциалы, t- ә сер потенциаланың ұ зақ тығ ы.
Ходжкин мен Хаксли мембранадағ ы кернеуді тұ рақ тандыру арқ ылы жү різген тә жірибелерін қ орытындылай келе, мембранадағ ы қ озу қ ұ былысының электрлік эквиваленті схемасын ұ сынды (4 сурет).
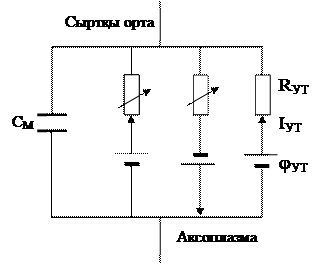 |
RK RNa
IK
jK jNa
IK
4 сурет
Бұ л модельге сә йкес қ озғ ан мембрана бір біріне параллель жалғ анғ ан С конденсатордан (мембраның сиымдылығ ына сә йкес), айнымалы екі кедергіден, олар калии gK = 1/RK жә не натрии gNa = 1/RNa каналдарының жә не ү шінші кедергі хлор жә не басқ а да иондардың gCl = 1/RCl ө ткізгіштігін сипатайды жә не осы тізбектегі тыныштық потенциалын сипаттайтын ү ш ЭҚ К тұ рады. Сонда қ озғ ан мембрана арқ ылы ө тетін ток мына шамамен сипатталады: IM = CM dj/dt + IK + INa + I, мұ ндағ ы CM dj/dt - сиымдылық тоғ ы (мембрананың қ айта зарядталуы кезіндегі ток шамсы), IK- калии иондарының тоғ ы, INa – натрии иондарының тогы, I – хлор жә не басқ а да иондар тоғ ы. Мұ ндағ ы ә р иондар тогы сол иондардың диффузиясы нә тижесінде пайда болатын тыныштық потенцила jМ мен тепе-тең дік Нернст потенциалнының jiP айрымына тең: Ii = gi(jM - jiP).
Осы айтылғ андарды ескере келе Ходжикин мен Хаксли ә сер потенциалы ү шін мына тү рдегі тең деуді ұ сынды:
IM = CM dj/dt + gK(jM - jKP) + gNa(jM - jNaP) + gУТ(jM - jУТP).
Бұ л ә рекет потенциалы ү шін Ходжкин-Хаксли тең деуі деп аталады жә не жү ргізілген эксперимент нә тижелерімен жақ сы ү йлеседі.
Жасуша мембранасының қ озу уақ ытының ұ зақ тығ ы ө те аз, 0, 5-1 мс аралығ ында жатады, онан соң мембрана қ абатының натрии ионын ө ткізуі тө мендейді, қ айтадан тыныштық потенциалы орнайды. Бұ л қ ұ былыс реполяризация деп аталады. Ә рекет потенциалының ұ зақ тығ ы жасуша тү ріне жә не температурағ а байланысты болады. Мысалы, температураны 100С кеміткенде ә рекет потенциалының ұ зақ тығ ы ү ш есе артады.
Қ озғ ан мембранадағ ы ә рекет потенциалы нерв талшығ ы (аксон) бойымен тарайды. Оның таралу механизмі тө мендегіше: мембрананың қ озғ ан бө лігінің ішкі қ абаты оң зарядталады (деполяризацияланады), ал қ озбағ ан бө лігінің заряды (тыныштық потенциалы) теріс қ алпында қ алады, екі бө лік арасында потенциалдар айырымы пайда болып, ток жү реді, ол ток ө ткізгіш сыртқ ы қ абат арқ ылы да жү ріп тізбекті тұ йық тайды (5- сурет).
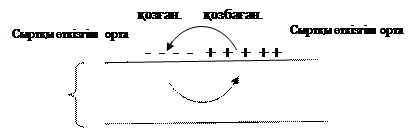
+ + + + - - - - - - - - -
аксон
+ + + + - - - - - - - - -
- - - - - + + + + + + +
5- сурет.
Осылайша пайда болғ ан «жергілікті (локальный) ток» деполяризацияланғ ан бө лікке жанасып жатқ ан қ озбағ ан нерв талшығ ының бө лігін тітіркендіріп, оны қ оздырады, нә тижесінде ол бө лікте ә рекет потенциалы пайда болады, бұ л қ ұ былыс келесі бө лікке ауысып, жалғ аса береді. Ток жасушаның сыртқ ы қ абаты арқ ылы қ озбағ ан бө ліктен қ озғ ан бө ліке қ арай жү реді, ал ішкі ортада ток керісінше бағ ытталады.
Кез келген электр тогы сияқ ты «жергілікті ток» мембрананы тіріркендіріп, мембрананың иондарды ө ткізуін жоғ арылатады, бұ л қ ұ былыс ө з кезегінде осы бө лікте ә рекет потенциалын тудырады, ол келесі бө лікті қ оздырады, процесс осылайша жалғ асып, нерв талшығ ы бойымен импульс тарайды. Бұ л кезде бұ рын қ озғ ан мембрана бө лігінде қ айта қ алпына келу процессі, яғ ни реполяризация (тыныштық потенциланының орнауы) қ ұ былысы жү реді. Импульстің аксон бойымен тарауы бір бағ ытта мембрананың қ озғ ан бө лігінен келесі бө лігіне қ арай жү реді. Мембрананың осының алдында ғ ана қ озуды ө ткізген бө лігі біраз уақ ыт бойы қ озбағ ан - рефракторлы кү йін сақ тайды, сондық тан импульс кері бағ ытта тарамайды.
Нерв импулісінің (ә рекет потенциалының) таралуы нерв талшығ ының қ асиетіне байланысты. Нерв талшығ ы миелинді жә не миелинсіз деген екі тү рге бө лінеді. Миелинді нерв талшығ ының цилиндр тә різді орта бө лігі аксоплазмамен толтырылғ ан, сырты цитоплазмалы мембранамен қ апталғ ан болып келеді. Нерв талшығ ының цилиндр тә різді ө сінің ә р жерінің 0, 2 мкм ден 2 мкм сайын қ абаты жаң алаштанғ ан болып келеді. Мұ ндай аймақ тарды нерв талшығ ының тү йіні немесе Ранвье ү зілісі деп атайды жә не бұ л аймақ тар жасушаның сыртын алып жатқ ан ертіндімен жанасады. Ранвье ү зіндісінің ұ зындығ ы жобамен 1 мкм тең (6- сурет).
Нерв жасаушасын миелинді қ абаты липидтер мен ақ уыздардан тұ рады сондық тан ол изолятор болып табылады. Осы себепті қ озу аксон мембранасының тек ашық аймақ тарында (Ранвье ү зіндісінде) пайда болады, бұ л кезде аксонның басқ а аймақ тарында потенциалдар айырымы тұ рақ ты, сыртқ ы орта оң, ішкі орта теріс зарядталғ ан болып келеді.
 |
6-cурет
Миелинді нерв қ абатынның деполяризацияланғ ан А бө лігіндегі қ озу келесі Б бө лігіне ө теді, бұ л қ ұ былыс ар қ арай тек В нү ктесінде жалғ асады, ал бұ рын қ озғ ан кү йде болғ ан Б нү ктесі қ озуды сезбейтін рефакторлы кү йде біраз уақ ыт болады. Осы себепті импульстер нерв арқ ылы тек бір бағ ытта ғ ана тарайды (7- сурет).
 |
7- сурет.
Миелинді нерв талшығ ы арқ ылы импульстердің осылайша таралуын секірмелі немесе сальтоторлы деп атайды. Импульстің сальтаторлы таралуы нерв импульстерінің таралу жылдамдығ ын артыруғ а мү мкіндік береді жә не мембрананың ө те аз бө лігінің қ озуы аз иондарды тасмалдауғ а, яғ ни натрии жә не калии иондық насостарының жұ мысына аз энергия жұ мсауғ а мү мкіндік туғ ызады.
Нерв талшығ ы арқ ылы ә сер потенциалының х қ ашық тық а жә не t уақ ытқ а байланысты таралуы телеграф тең деуі арқ ылы сипатталады екен жә не оның шешуі мына ө рнекке тең:
j = j0е-х/l,
мұ ндағ ы j - х нү ктедегі потенциал мә ні, х- қ озғ ан жерден потенциалы анық талынатын нү ктеге дейінгі қ ашық тық, l - нерв талшығ ының тұ рақ тысы, ол деполяризия потенциалының е есе кемуіне тең ара қ ашық тық. Нерв талшығ ыынң тұ рақ тысы мына шамағ а тең:
l = Ö rMDa/2ri,
мұ ндағ ы rM- нерв талшығ ы қ абатының меншікті электр кедергісі, D - қ абат қ алың дығ ы, a – нерв талшығ ының радиусы, ri – цитоплазманың меншікті электр кедергісі. Бұ л ө рнектерден миелинсіз нерв талшығ ы ү шін нерв талшығ ы тұ рақ тысының ү лкен шама болғ ан сайын импульстің таралу жылдамдығ ы артатындығ ы байқ алады, ал ол ө з кезегінде аксон радиусы мен мембрананың меншікті кедергісінің ү лкен, цитоплазманың меншікті кедергісінің аз болғ андығ ын сайын ү лкен шама болатындығ ын кө реміз.
Миелинді нерв талшығ ында қ озудың таралуы Ранвье ү зіндісінің ұ зындығ ына байланысты болады екен. Жү ргізілген ө лшеулер импульстің таралуы бір ү зілістен екінші ү зіліске дейін 0, 07 мс екендігін кө рсетті. Нерв радиусы 10 мкм болатын талшық тағ ы импульстардың таралу жылдамдығ ы 120 м/с тең.
Аутоиммундық аурулардың бірі склероз нерв талшығ ының миеленді қ абаттың бұ зылуынан, нерв талшығ ының ашылып қ алуынан пайда болады. Соның ә серінен нерв импульстарының ашылып қ алғ ан нерв аймағ ындағ ы таралу жылдамдығ ы нашарлайды, нә тижесінде кө ру мен қ озғ алу қ абілеті тө мендейді, бұ лшық еттердің ә лсізденуі байқ алады т.б. Сондай ақ - нерв талшығ ының демиелендігі басқ а да аурулар себебінен де пайда болуы мү мкін, мысалы невралгия, радикулит, тү рлі полиневропатология.
|
|