Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Нейронные механизмы научения
|
|
В основе процессов научения и памяти, проявляющихся на поведенческом уровне целостного организма, лежит свойство пластичности нейрона, т.е. способности его к перестройке своих функциональных свойств. Выделяют несколько основных проявлений пластичности: 1) привыкание; 2) сенситизация; 3) клеточные аналоги ассоциативного обучения; 4) долговременная посттетаническая потенциация; 5) долговременная депрессия; 6) пластичность пейсмекерного механизма клетки.
Привыкание нейрона выражается в постепенном ослаблении его реакции на повторяющийся относительно нейтральный раздражитель. Если же параметры раздражителя изменяются или появляется новый стимул, то реакция нейрона восстанавливается.
Сенситизация (сенсибилизация) нейрона – это временное усиление его реакции или появление ее на ранее неэффективный раздражитель, особенно в тех случаях, когда он неоднократно связывается с субъективно приятными или неприятными воздействиями. Сенситизация по значению противоположна привыканию. Например, сенситизация наступает при переходе из света в темноту в результате темновой адаптации, когда сначала не видимые предметы после некоторого нахождения в темноте начинают восприниматься палочками сетчатки. Сенситизация может сохраняться от нескольких секунд до дней и недель. Считают, что сенситизация обусловлена активацией модулирующих нейронов, вызываемой сильным посторонним раздражителем.
Клеточные аналоги ассоциативного обучения можно наблюдать у нейронов многих структур мозга. Примерно 40 % нейронов гиппокампа способны к обучению в виде формирования условного рефлекса. Условный рефлекс образуется у нейрона в том случае, если к нему последовательно, через небольшое время поступают сигналы от условного и безусловного раздражителей. Условным стимулом может стать любое, вначале индифферентное сенсорное или электрическое раздражение, которое вызывает лишь подпороговую реакцию в виде ВПСП. Безусловный раздражитель должен вызывать у нейрона разряды, т.е. потенциалы действия. Под влиянием неоднократного сочетания индифферентного раздражителя с безусловным нейрон, способный к ассоциативному обучению, начинает отвечать на ставший условным (бывший индифферентный) стимул реакцией, которая ранее возникала только на безусловный раздражитель.
 В осуществлении привыкания, сенситизации и ассоциативного обучения нейрона определяющую роль играют ионные механизмы синаптической передачи нервных импульсов, сильно зависящие от активности модулирующих нейронов. При развитии привыкания модулирующие нейроны не активны. Многократно повторяющееся поступление одинаковых сигналов в пресинаптический отдел вызывает инактивацию кальциевых каналов в нервном окончании. Поступление ионов Ca2+ в пресинаптический отдел падает и количество выделяемого в синаптическую щель медиатора уменьшается (рис. 106, А).
В осуществлении привыкания, сенситизации и ассоциативного обучения нейрона определяющую роль играют ионные механизмы синаптической передачи нервных импульсов, сильно зависящие от активности модулирующих нейронов. При развитии привыкания модулирующие нейроны не активны. Многократно повторяющееся поступление одинаковых сигналов в пресинаптический отдел вызывает инактивацию кальциевых каналов в нервном окончании. Поступление ионов Ca2+ в пресинаптический отдел падает и количество выделяемого в синаптическую щель медиатора уменьшается (рис. 106, А).
При сенситизации, вызываемой сильным посторонним стимулом, который действует одновременно с основным раздражителем, активируются модулирующие нейроны. Они образуют на пресинаптических окончаниях аксона чувствительного нейрона синапсы, выделяющие медиатор серотонин. Серотонин взаимодействует с аденилатциклазой, что вызывает инактивацию калиевых каналов, приводящую к удлинению потенциала действия у чувствительного нейрона. В результате чувствительный нейрон выделяет больше, чем обычно, медиатора в синаптическую щель и реакция постсинаптического нейрона возрастает (рис. 106, Б).
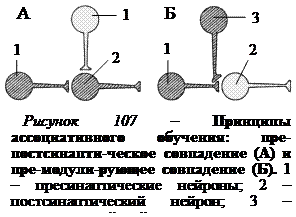
При ассоциативном обучении процессы, происходящие в нейронах, более сложны, чем во время привыкания и сенситизации. Особенностью ассоциативного обучения является наличие подкрепления, которым может являться сильное безусловное раздражение. Было выдвинуто два принципа ассоциативного обучения. Согласно принципу пре-постсинаптического совпадения, предложенного Д. Хеббом, необходимо совпадение активности пресинаптического и постсинаптического нейрона для усиления связи между ними (рис. 107, А). Э. Кэндел и Р. Хокинс выдвинули принцип пре-модулирующего совпадения, или зависящего от активности обучения.
В случае пре-модулирующего совпадения на пресинаптическом окончании сенсорного нейрона расположен серотонинергический синапс модулирующего нейрона (рис. 107, Б). В результате поступления импульсов в пресинаптическое окончание по аксону сенсорного и модулирующего нейронов возникает эффект, аналогичный сенситизации, т.е. происходит облегчение выделения медиатора и передачи импульса к постсинаптическому нейрону.
 Долговременная посттетаническая потенциация (синаптическое облегчение, постсинаптическая потенциация) – это улучшение проведения в синапсах после кратковременного раздражения афферентных путей. Впервые ее обнаружил у нейронов гиппокампа норвежский ученый П. Андерсен с сотрудниками. Применив высокочастотную электрическую стимуляцию для вызывания короткой серии потенциалов действия в пресинаптическом нейроне, они регистрировали усиление ответов постсинаптического нейрона на последующие одиночные импульсы, приходящие от пресинаптической клетки. Было установлено, что долговременная потенциация обусловлена усиленным выделением медиатора из пресинаптических окончаний.
Долговременная посттетаническая потенциация (синаптическое облегчение, постсинаптическая потенциация) – это улучшение проведения в синапсах после кратковременного раздражения афферентных путей. Впервые ее обнаружил у нейронов гиппокампа норвежский ученый П. Андерсен с сотрудниками. Применив высокочастотную электрическую стимуляцию для вызывания короткой серии потенциалов действия в пресинаптическом нейроне, они регистрировали усиление ответов постсинаптического нейрона на последующие одиночные импульсы, приходящие от пресинаптической клетки. Было установлено, что долговременная потенциация обусловлена усиленным выделением медиатора из пресинаптических окончаний.
Это выделение стимулируется специальным веществом (возможно, окисью азота), которое образуется в постсинаптическом нейроне и поступает ретроградно через синаптическую щель в пресинаптический нейрон. Синтез этого вещества осуществляется через цепь реакций, которые запускаются при избытке ионов кальция, поступающих в постсинаптический нейрон через каналы, связанные с NMDA-рецепторами (рис. 108). Рецепторы получили такое название, поскольку активируются синтетическим аналогом глутамата N-метил-D-аспартатом. NMDA-рецепторы локализованы в постсинаптической мембране, медиаторами для них служит глутамат. Ворота каналов, связанных с NMDA-рецепторами, открываются только в том случае, когда постсинаптическая мембрана сильно деполяризована, а к рецепторам присоединен медиатор глутамат. Деполяризация постсинаптической мембраны снимает блокаду кальциевых каналов в NMDA-рецепторах, которая осуществляется внеклеточным магнием. В постсинаптической мембране, кроме NMDA-рецепторов, имеются так-же не-NMDA-рецепторы, которые под действием глутамата сразу открывают кальциевые ионные каналы даже в отсутствие сильной деполяризации мембраны.
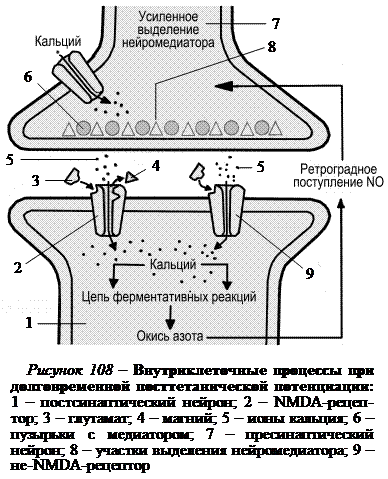 Каналы, связанные с NMDA-рецепторами, находясь в открытом состоянии, пропускают очень большой поток ионов Ca2+. NMDA-рецепторы играют основную роль в долговременной посттетанической потенциации. Если их избирательно заблокировать, то долговременная потенциация не происходит, холя синаптическая передача не нарушается. Искусственно регулируя внутриклеточный уровень ионов Ca2+, можно усилить долговременную потенциацию или предотвратить ее.
Каналы, связанные с NMDA-рецепторами, находясь в открытом состоянии, пропускают очень большой поток ионов Ca2+. NMDA-рецепторы играют основную роль в долговременной посттетанической потенциации. Если их избирательно заблокировать, то долговременная потенциация не происходит, холя синаптическая передача не нарушается. Искусственно регулируя внутриклеточный уровень ионов Ca2+, можно усилить долговременную потенциацию или предотвратить ее.
Степень выраженности синаптического облегчения возрастает с увеличением частоты импульсов. Оно максимально, когда импульсы поступают в синапс с интервалом в несколько миллисекунд. Электронно-микроскопическими исследованиями подтверждена связь долговременной потенциации с увеличением количества синапсов на дендритах нейронов гиппокампа и изменением структуры синапсов. Долговременная постсинаптическая потенциация выявлена и в других структурах мозга: мозжечке, миндалине.
Длительность посттетанической потенциации зависит от свойств синапса и характера раздражения. После одиночных стимулов она выражена слабо, но после серии раздражений посттетаническая потенциация может продолжаться от нескольких минут до нескольких часов. Это крайне важно как для выработки условных рефлексов, так и для любого обучения.
Долговременная депрессия (синаптическая депрессия) была открыта в мозжечке у клеток Пуркинье, а впоследствии обнаружена в гиппокампе, таламусе, коре больших полушарий и др. Она заключается в длительном снижении проводимости через синапс и развивается при продолжительном поступлении импульсов. Долговременную депрессию можно вызвать совместным низкочастотным раздражением двух входов к клетке Пуркинье: входа к ее дендритам через параллельные волокна и входа от лазающих волокон. Долговременная депрессия объясняется расходованием медиатора, накоплением продуктов распада веществ, в частности повышением кислотности среды при длительном проведении возбуждения по одним и тем же нейронным цепям.
Пластичность пейсмекерного механизма – это способность особых нейронов-пейсмекеров изменять свою активность под влиянием поступающих сигналов, вызываемых последовательными раздражениями. Пейсмекеры (водители ритма) представляют собой источники активности мозга, генерирующие собственный эндогенный ритм. В роли пейсмекеров могут выступать нейронная сеть или отдельные нейроны, способные генерировать ритмическую импульсную активность.
Самый простой сетевой пейсмекер состоит из двух нейронов, соединенных положительной обратной связью. Такая структура обеспечивает межклеточную реверберацию сигнала (его многократное отражение, повторение движения по кругу). Воздействуя на один из нейронов сетевого пейсмекера, можно управлять его работой, включая или выключая реверберацию нервного импульса.
Пейсмекером может быть и один нейрон, в котором возникает внутриклеточная реверберация импульса от тела нейрона к окончанию аксона и обратно или в сому (тело) которого встроен генератор ритмической активности. Синаптические и гуморальные влияния, действие пептидов, внесинаптическое влияние медиаторов включают и выключают режим пейсмекерной активности.
Пейсмекерные нейроны используются для управления реакциями организма. С их помощью реализуются генетические программы, например, локомоции (ходьба, бег).
Пейсмекерный механизм обладает пластичностью, которая выражается в том, что ответ нейрона меняется по мере повторения действия одиночных раздражителей. В том случае, когда ответ пейсмекерного нейрона слабеет от раздражения к раздражению, этот процесс по аналогии с угасанием поведенческой реакции называют привыканием. Если реакция нейрона от раздражения к раздражению возрастает, то это называется фасилитацией.
Пластические свойства пейсмекерного механизма существенно расширяют адаптационные возможности организма в использовании наследственных (генетических) программ поведения.
|
|