Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Обмен метаболитами ЦТК между митохондриями и цитозолем
|
|
Цикл трикарбоновых кислот — это не только необходимый этап энергообмена, но и источник соединений, необходимых для многих биосинтезов, протекающих в цитозоле и других компартментах. Благодаря интенсивному обмену с цитозолем в митохондриях пересекается обмен трех групп важнейших соединений — углеводов, белков и липидов (рис. 4.8).
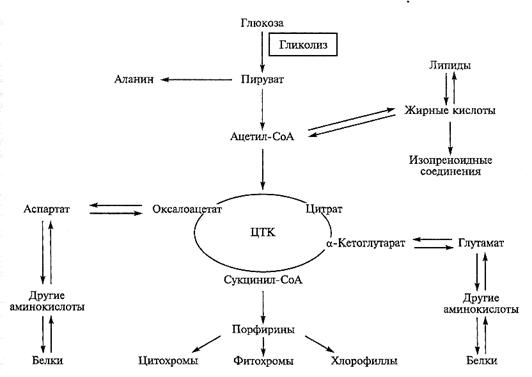
Рис. 4.8. Цикл трикарбоновых кислот как центр метаболизма.
Интермедиаты ЦТК активно используются как исходные субстраты при образовании аминокислот, липидов и во многих других биосинтезах. Предлагаемая схема не является полной, она лишь демонстрирует активный отток метаболитов с основного пути дыхания
Например, ацетил-СоА — исходное соединение для синтеза жирных кислот и полиизопреноидных соединений, необходимых при биосинтезе липидов, каротиноидов, растительных гормонов. Важнейшей функцией цикла является экспорт так называемых «углеродных скелетов» в виде кетокислот, необходимых для синтеза аминокислот в реакциях переаминирования.
Интеграция митохондрий в общий метаболизм возможна благодаря интенсивному обмену метаболитами между матриксом и цитозолем. Наружная мембрана благодаря наличию пор не препятствует такому обмену. Транспорт соединений через внутреннюю мембрану осуществляется с помощью интегрированных в нее белков-транспортеров. Транспортеры переносят кето- и аминокислоты, адениннуклеотиды, НАД+, коэнзим А и многие другие соединения (см. также подразд. 4.2.11). Так, во внутренней мембране есть целое семейство похожих по своей структуре белков, которые осуществляют транспорт анионов ди- и трикарбоновых кислот, участвующих в ЦТК
(рис. 4.9). Особенно интенсивно из митохондрий идет вынос α -кетоглутарата, который необходим для ассимиляции аммония и синтеза аминокислот в хлоропластах. В мембране обнаружены два обменных переносчика, которые выносят в цитозоль α -кетоглутарат и/или цитрат в обмен на малат. В цитозоле цитрат превращается в изоцитрат (при участии аконитазы), а затем окисляется с образованием α -кетоглутарата в реакции, катализируемой НАДФ+-зависимой изоцитратдегидроге-назой. В обмен на а-кетоглутарат или цитрат в митохондрии поступает малат, тем самым возмещая углеродные потери ЦТК.
Между митохондриями и цитозолем возможен также обмен восстановительными эквивалентами. Образующийся в ЦТК НАДН не только окисляется в дыхательной цепи, но и выносится в цитозоль, где используется как восстановитель в разных реакциях, например при восстановлении нитрата (см. гл. 6). И, наоборот, в ряде случаев НАДН из цитозоля может доставляться в матрикс. В то же время белок-переносчик для НАДН неизвестен. Обмен НАДН между матриксом и цитозолем осуществляется с помощью так называемых «челночных систем», или «шаттлов». Наиболее важная из них — малат/оксалоацетат-шаттл (рис. 4.9), оперирующий при участии
НАД-зависимой малатдегидрогеназы, изоформы которой есть и в матриксе, и в цитозоле. В митохондриях, где содержание НАДН высокое, оксалоацетат за счет НАДН восстанавливается до малата. В мембране присутствует белок-переносчик, обменивающий малат на оксалоацетат. Оказавшись в цитозоле, малат может быть вновь окислен в обратной реакции с образованием НАДН. Этот простой челнок в принципе может работать в двух направлениях в зависимости от соотношения НАДН/НАД+ по обе стороны митохондриальной мембраны. В то же время есть данные, что in vivo он работает в основном «на экспорт»: около 25 —50 % НАДН, образованного в матриксе, окисляется в цитозоле благодаря малат/оксалоацетат-шаттлу. Цикл трикарбоновых кислот не только поставляет метаболиты, необходимые для синтеза белков и липидов, но также включен в деградацию этих соединений. Хотя растения «дышат» в основном углеводами, в некоторых случаях дыхание поддерживают белки или жиры. Например, при развитии проростков образованные при распаде запасных белков аминокислоты через реакции переаминирования превращаются в кетокислоты — интермедиаты ЦТК и включаются в цикл. Использование жиров при прорастании семян масличных растений представляет собой особый случай, связанный с функционированием глиоксилатного цикла.
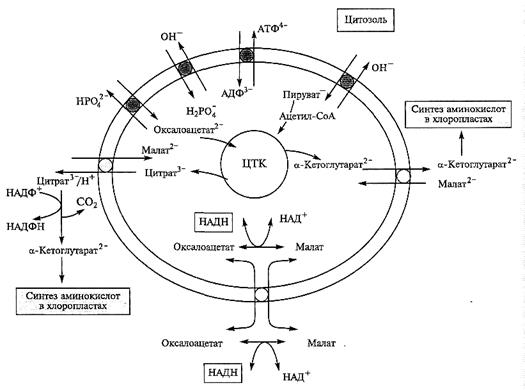
Рис. 4.9. Обмен метаболитами между матриксом и цитозолем.
Внутренняя мембрана митохондрий содержит белки-переносчики. Пируват и оксалоацетат поступают в митохондрии в обмен на ион ОН  или фосфат и метаболизируют в ЦТК с образованием цитрата. Вынос цитрата или α -кетоглутарата в цитозоль в обмен на малат обеспечивает углеродными скелетами синтез аминокислот в пластидах, в том числе в хлоропластах. В цитозоле цитрат может превращаться в α -кетоглутарат при участии цитозольной НАДФ-зависимой изо-цитратдегидрогеназы. Малат/оксалоацетат шаттл обеспечивает обмен НАДН между матриксом и цитозолем. Шаттл работает благодаря переносчику (известному как ОАА-транспортер), который осуществляет обменный транспорт через мембрану малата и оксалоацетата в ту и другую сторону. Важным транспортером является АТФ/АДФ-транслокатор и переносчик фосфата (см. подразд. 4.2.11)
или фосфат и метаболизируют в ЦТК с образованием цитрата. Вынос цитрата или α -кетоглутарата в цитозоль в обмен на малат обеспечивает углеродными скелетами синтез аминокислот в пластидах, в том числе в хлоропластах. В цитозоле цитрат может превращаться в α -кетоглутарат при участии цитозольной НАДФ-зависимой изо-цитратдегидрогеназы. Малат/оксалоацетат шаттл обеспечивает обмен НАДН между матриксом и цитозолем. Шаттл работает благодаря переносчику (известному как ОАА-транспортер), который осуществляет обменный транспорт через мембрану малата и оксалоацетата в ту и другую сторону. Важным транспортером является АТФ/АДФ-транслокатор и переносчик фосфата (см. подразд. 4.2.11)
|
|