Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Психомоторную деятельность
|
|
Формирование представлений о мозговых механизмах произвольных движений, изначально базировавшихся на понятии о " двигательных центрах" коры, проделало значительный исторический путь, каждый этап которого связан главным образом, с одной стороны, с научными открытиями в области морфологии и физиологии мозга и, с другой стороны, с теми научными концепциями и взглядами ученых, в контексте которых эти открытия трактовались. Так, к примеру, складывавшиеся к концу XIX-го века представления о том, что постцентральная область мозговой коры является зоной чувствительных проекций, в то время как центральная извилина представляет собой двигательный аппарат коры головного мозга, регулирующий произвольные движения путем посыла импульсов по эфферентному пирамидному пути к передним рогам спинного мозга, в дальнейшем благодаря рефлекторной концепции движений, сформулированной И.П. Павловым и его учениками (Н И. Красногорским, 1911, Ю.М. Конорским и С.М Миллером, 1936 и др.) и разработанной позже Л.А. Орбели (1935). П.К Анохиным (1935) и Н.А. Бернштейном (1947), претерпели значительные изменения.
Благодаря основным положениям павловской рефлекторной концепции движений о ведущей роли в организации моторных актов системы афферентных синтезов, формирующих двигательные импульсы на выполнение этих моторных актов, происходит замена понятия о " двигательных центрах" коры более широким понятием " корковых отделов двигательного анализатора" Согласно этому представлению, в корковые отделы двигательного анализатора входит большой комплекс корковых зон, которые обеспечивают ту или иную сторону подготовки двигательного акта. Так, отмечает А.Р. Лурия (1962), одни зоны осуществляют пространственную организацию произвольных движений, другие анализируют импульсы от мышечно-суставного аппарата, наконец, третьи обеспечивают направляющее и регулирующее влияние речевых связей, которое, как он подчеркивает, является " важнейшим составным элементом организации произвольного движения" (А.Р. Лурия, 1962; С. 149; выделено нами - В.Д.). По мнению Н.А. Бернштейна (1947), А.Р. Лурия (1957) и других авторов, все эти области мозга, каждая из которых вносит свой компонент в организацию моторного акта, обеспечивая ту или иную сторону построения движения, должны быть включены в состав корковых отделов двигательного анализатора в широком смысле этого понятия.
В узком смысле данного понятия представление о корковых отделах двигательного анализатора связано с выделением " коркового ядра", или " ядерной зоны" двигательного анализатора. Термины " ядерная зона" и " анализатор", понятия о них, а также представления о системах анализаторов и ядерных зонах в коре головного мозга впервые были даны в работах И.П. Павлова (1949). Под ядерными зонами, как известно, И.П. Павлов понимал определенные области коры, в которых имеется максимально выраженная концентрация специфических элементов соответствующих анализаторов вместе с их связями. " Ядерные" элементы этих зон, по мнению И.П. Павлова, тесно сплетены между собой и выполняют самые сложные анализы и соответственно синтезы специфических для деятельности отдельных анализаторов раздражений.
Особенностью коркового ядра двигательного анализатора, отличающей его от соответствующих отделов других анализаторов, является то, что к нему, как указывает А.Р. Лурия, " можно отнести не одну, а две совместно работающие зоны мозговой коры, которые обычно обозначаются как постцентральная и прецентральная области и которые вместе образуют единый функциональный аппарат - " сензомо-торную область коры (1962; С. 149; выделено — А.Л.).
Формирование начальных представлений о механизме движений стало возможным после того как в 70-е годы Х1Х-то века на основе проведенных экспериментов G. Fnsch u. E. Hitzig (1870) выделили спереди от центральной борозды двигательную область коры — изолированные " двигательные центры". В последующих исследованиях ученых эта область мозга приобретала все более четкую дифференцированную локализацию, обнаруживаемую в опытах на выявление связи между раздражаемыми участками мозга и сокращениями различных мышц и мышечных групп. Так, были обнаружены в верхней части передней центральной извилины центры движения стопы, голени и бедра, ниже в этой же извилине - клеточные группы, связанные с движениями предплечья, пальцев руки, еще ниже - центры движения мышц лица, языка, челюстей, мягкого неба и гортани.
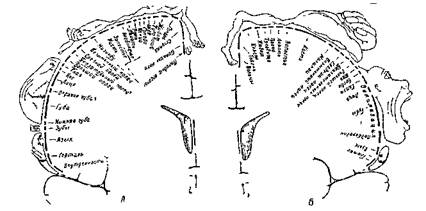
Сложившиеся на сегодня представления о том, что передняя центральная извилина являет собой первичную двигательную зону со cтрогo определенной проекцией участков тела, являются общеизвестными и общепризнанными. Установлено, что лицо и органы артикуляции спроецированы, как было уже отмечено, в нижней примерно трети указанной извилины, кисть руки и пальцы - в средней трети, а туловище, живот и таз представлены в верхней трети этой же извилины, двигательная же область ноги - на медиальной (срединной) ее поверхности. Таким образом, тело человека относительно его двигательной системы спроецировано в передней центральной извилине вверх ногами и соответственно вниз головой. Результаты зарубежных исследований угой направленности, провидимых в 50-е годы 20-го столетия, наглядно представлены широко известной сегодня и в отечественной специальной литературе схемой W. Penfield (1959), иллюстрирующей данные его многочисленных экспериментов (рис.1).
Рис.1. Схема соматотопической проекции в коре головного мозга (по W.Penfield, 1959)
А корковая проекция чувствительности, Б - корковая проекция двигательной системы (Относительные размеры органов отражают ту площадь коры головного мозга, с которой могут быть вызваны соответствующие ощущения и движения)
Довольно продолжительное время с начала первых исследований в области движений человека никакой физиологической теории о работе двигательной зоны коры не существовало. Более того, еще несколько десятилетий после указанного выше открытия G. Fnsch u. Е Hitzig (1870), как известно из истории физиологии, ученые многих
стран вели споры о том, считать ли переднюю центральную извилину подлинно двигательной зоной или рассматривать ее как чувствительную, но связанную с двигательными центрами.
Дальнейшее развитие представлений о двигательном отделе коры стало возможным благодаря разработанной И.П. Павловым теории условных рефлексов, раскрывавшей физиологические механизмы работы коры головного мозга. Так, уже в 1911 году проведенными учеником И.П. Павлова Н.И. Красногорским опытами на собаках с использованием метода условных рефлексов была впервые выявлена возможность выработки двигательного условного рефлекса.
Этими опытами, как отмечает Е.И. Бойко (1969), были доказаны два теоретических положения:
" 1) что двигательная область коры, строго говоря, не является двигательным центром, а афферентной, чувствующей областью, принимающей с периферии сигналы о движении мышц, и
2) что поля 4 и 6 коры (рис. 2) являются двигательными анализаторами, подобно всем другим известным анализаторным зонам коры.. " (Е.И. Бойко, 1969; С.48). Благодаря этим положениям был сделан вывод о том, что функция двигательного анализатора состоит в физиологическом различении разнообразных движений конечностей и всех мышц тела и последующем связывании их с двигательными центрами спинного мозга. Этот вывод Н.И. Красногорского и И.П. Павлова был высказан последним довольно определенно: " Красногорский... точно установил несомненную афферентную природу двигательной области коры" (И.П. Павлов, 1949, С.553).
Такой точки зрения вплоть до 60-70-х годов 20-го века придерживались многие исследователи. Так, к примеру, Е.И. Бойко отмечает, что " двигательная зона по существу не двигательная, а чувствующая, афферентная, такая же в сущности, как и кожная, но только прямо связанная через пирамидный путь с двигательными центрами спинного мозга" (1969; С.49). Более того, замечает он, " неврология приносит все новые и новые данные, доказывающие, что поля 4 и 6 коры получают по специальным волокнам раздражения от рецепторов, расположенных в мышцах и суставах, а поля 1, 2 и 3, принимая в себя часть этих импульсов, по другим путям получают гораздо более мощные афферентные импульсы от кожных рецепторов". По мнению этого автора, " оба анализатора - двигательный и кожный", расположенные соответственно в передней и задней центральных извилинах, " имеют различные нервные механизмы, хотя и работают, как правило, совместно" (Е.И. Бойко, 1969; С.49-50; выделено - Е.Б.). Подчеркивая заслугу Н.И. Красногорского в разделении работы этих двух систем (двигательной и кожной) и тем самым в доказательстве существования самостоятельного двигательного анализатора в передней центральной извилине Е И Бойко, как следует из его рассуждений, придерживается этой же точки зрения.
С такой позицией, ее несколько односторонним обоснованием и соответствующими выводами едва ли можно согласиться, принимая во внимание ряд следующих принципиальных моментов.э Во-первых, еще с павловских времен, как известно, двигательная область коры в передней центральной извилине продолжала рассматриваться целым рядом исследователей именно как двигательная, а не афферентная, чувствительная Во-вторых, как раз в 60-70 е годы прошлого столетия, когда, по мнению Е И Бойко были доказаны с помощью данных неврологии выводы И. П. Павлова и Н И Красногорского, проводились многочисленные анатомические, физиологические, клинические и другие исследования мозга (Л.А. Кукуев, М.М. Кольцова, А.Р. Лурия, Г.И. Поляков И.Н. Филимонов и др.), благодаря которым были получены новые важные данные о структурно-функциональной организации двигательного анализатора, далеко не во всем подтверждающие приведенные выше выводы И.П. Павлова и Н.И. Красногорского. Кроме того, исследованиями именно этого периода были значительно обогащены представления о роли двигательной коры в целостной интегративнои деятельности мозга, а также выявлен ряд других мозговых образовании, принимающих участие в организации психомоторной деятельности человека
Ознакомление с основными исследованиями, явившимися итогом плодотворного изучения мозговой организации движении животных и человека за более чем полувековой период времени, а также более обстоятельный анализ отечественной и зарубежной литературы по данной проблематике, вышедшей за последние 10-15 лет, позволяют составить вполне определенное представление об историческом развитии и сегодняшнем состоянии этой области знаний и вместе с тем получить необходимые в контексте рассматриваемого вопроса сведения о морфологических и функциональных особенностях двигательного анализатора как главного аппарата в механизме психомоторных проявлений человека. Эти знания и составляют сегодня ту естественно научную и методологическую базу на которой выстраиваются современные исследования в данной области научного познания.
Основу современных представлений о мозговой организации психомоторной деятельности человека составляют, прежде всего, имеющиеся на сегодня научные данные об участии отдельных образований нервной системы в осуществлении двигательных актов и их роли в целостной интегративной работе мозга

Рис.2. Карта цитоархитектонических полей коры головного мозга (по данным Московского института мозга)
А) конвекситальная поверхность коры б) медиальная (срединная) поверхность
коры (Цифрами обозначены поля.
Сплошными линиями помечены границы полей, пунктирными подразделения внутри поля)
Исследованиями доказано, что " ядро" двигательного анализатора, базируется в двух центральных извилинах: передней (поля 4, 6) и задней (поля 1, 3), где осуществляются функции высшего анализа и синтеза двигательных раздражений. Как видно, и это необходимо подчеркнуть еще раз, что двигательный анализатор представлен одновременно передними отделами коры (моторная зона в передней цен-тральной извилине) и задними отделами коры (кожно-кинестетическая зона в задней центральной извилине), которые характеризуются существенно отличными структурными и функциональными особенностями, а также закономерностями взаимодействия с другими мозговыми образованиями.
Очевидно, что без знания особенностей строения и учета закономерностей функционального взаимодействия различных корковых отделов мозга, принимающих участие в организации психомоторных актов, не может получить научное обоснование и быть эффективной система работы по формированию психомоторики. Не могут быть также объективно выявлены и верно квалифицированы двигательные отношения, установлены их место и роль в структуре дефекта, правильно определены уровень и специфика психомоторного недоразвития в целом. Без этих знаний не могут быть также вскрыты причины и механизмы двигательных нарушений, степень их включенности и влияния на межсистемные функциональные расстройства и т.д., что в свою очередь не позволит обеспечить в каждом конкретном случае адекватность коррекционно-педагогических мероприятий.
Сказанное определяет значимость и необходимость рассмотрения морфологической и физиологической (психофизиологической) организации психомоторики при ее нормальном и патологическом функционировании. Основываясь на литературных данных, представляем в нижеследующих разделах морфофункциональные особенности упомянутых выше передних и задних отделов коры, входящих в состав двигательногo анализатора и обеспечивающих психомоторную деятельность человека.
|
|