Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Процессы торможения в нейронных цепях и сетях
|
|
Интегративная и координационная деятельность центральных нервных образований осуществляется при обязательном участии тормозных процессов. Торможение в ЦНС – это активный процесс, внешне проявляющийся в подавлении, замедлении или ослаблении процесса возбуждения и характеризующийся определенной интенсивностью и длительностью. Торможение нельзя свести ни к утомлению нервных центров, ни к их перевозбуждению, ни к катодической депрессии нервных клеток. Торможение в норме неразрывно связано с возбуждением, сопутствует возбудительному процессу, ограничивая и препятствуя чрезмерному его распространению. При этом торможение часто ограничивает возбуждение и вместе с ним формирует мозаику активированных и заторможенных зон в центральных мозговых структурах.
Явление торможения в нервной системе было открыто И.М.Сеченовым в 1862 году в виде эффекта центрального торможения. Суть его заключалась в том, что химическое раздражение кристалликом соли зрительных бугров (таламуса) лягушки вызывает торможение простых спинномозговых рефлекторных реакций.
Торможение – активный нервный процесс, приводящий к угнетению или предупреждению возбуждения. В зависимости от локализации тормозного процесса различают периферическое торможение, осуществляемое непосредственно в синапсах на мышечных и железистых элементах, и центральное, реализуемое в пределах центральной нервной системы.
Если торможение развивается как следствие возбуждения соответствующих дополнительных тормозящих или возбуждающих нейронов – это вторичное торможение.
Направленность эффекта (возбуждение или торможение) определяется изменением ионной проницаемости постсинаптической мембраны при взаимодействии медиаторов с рецепторами. Поэтому некоторые медиаторы способны опосредовать как возбуждение, так и Т.
У некоторых типов нейронов тормозные синапсы локализуются на телах и близких к ним участках Дендритов, что вследствие соседства с триггерной зоной генерации распространяющегося возбуждения обеспечивает высокую эффективность Т. Из этого правила есть исключения (например, тормозные синапсы звездчатых нейронов на клетках Пуркине мозжечка расположены на удалённых участках дендритов).
Функциональная значимость постсинаптического Т. разнообразна.
Афферентное (прямое) Т. служит для ослабления возбуждения функционально антагонистических элементов и тем самым способствует координированному, пространственно направленному протеканию возбуждения в цепях нейронов. В спинном мозге, в частности, такое Т. является основой так называемого реципрокного (взаимообратного) Т. мотонейронов, иннервирующих мышцы-антагонисты (см. Реципрокная иннервация). Возвратное (коллатеральное) Т., осуществляемое через систему возвратных коллатералей (ветвей) аксонов эфферентных нейронов и специализированных вставочных тормозных нейронов, стабилизирует собственный уровень возбуждения определённого структурно-функционального объединения (блока) нейронов и ограничивает распространение возбуждения на соседние популяции нейронов.
Менее изучено так называемое пресинаптическое Т., выражающееся в угнетении возбуждения в нервных терминалиях, то есть на входе постсинаптического клеточного элемента. Это Т. имеет необычайно большую длительность (сотни мсек) и совпадает во времени с проявлением деполяризации приходящих афферентов. Предполагают, что на деполяризации основано пресинаптическое Т., а его морфологическим субстратом являются аксо-аксональные синапсы, происхождение пресинаптических элементов которых неизвестно. Имеются веские аргументы в пользу роли гамма-аминомасляной кислоты как медиатора пресинаптического Т., по крайней мере в нервно-мышечных соединениях ракообразных и в спинном мозге позвоночных. По-видимому, Сеченовское торможение у лягушки осуществляется по механизму преспнаптического Т.
Известно также пессимальное, или вторичное, Т., выражающееся в блокировании возбуждения вследствие его чрезмерности (см. Парабиоз). Этот феномен, описанный впервые Н. Е. Введенским (См. Введенский), трудно выявить при физиологических условиях эксперимента, но можно демонстрировать при аномальных (в частности, судорожных) состояниях.
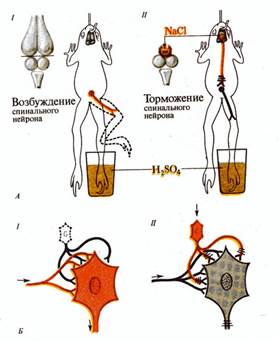
Рис. Сеченовское торможение.
Торможение в ЦНС можно классифицировать по различным признакам:
• по электрическому состоянию мембраны - деполяризационное и гиперполяризационное;
• по отношению к синапсу - пресинаптическое и постсинаптическое;
• по нейрональной организации – первичное и вторичное;
· по форме – латеральное (боковое), возвратное, реципрокное.
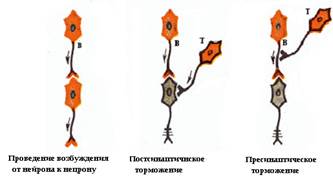

Рис. Проведение возбуждения от нейрона к нейрону. Локализация торможения – пресинаптическое и постсинаптическое. Пессимальное торможение
Первичное торможение обусловлено наличием специфических тормозных синапсов и развивается первично без предварительного возбуждения.
Вторичное торможение возникает без участия специализированных тормозных структур как следствие избыточной активации возбуждающих входов нейрона. Эту форму торможения определяют как торможение Введенского (пессимум), который открыл его в 1886 г. при исследовании нервно-мышечного препарата.
Торможение Введенского играет предохранительную роль и возникает при чрезмерной активации центральных нейронов в полисинаптических рефлекторных дугах. Оно выражается в стойкой деполяризации клеточной мембраны, превышающей критический уровень и вызывающей инактивацию Na+–каналов, ответственных за генерацию потенциалов действия.
Таким образом, процессы торможения в локальных нейронных сетях уменьшают избыточную активность и участвуют в поддержании оптимальных режимов импульсной активности нервных клеток.
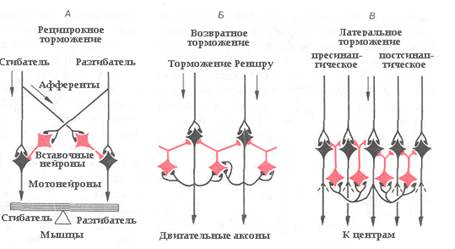
Рис. Формы торможения в ЦНС.
В нервных центрах могут быть как различные виды торможения, так и разные их сочетания. В результате различают следующие формы торможения. Возвратное торможение заключается в том, что аксон тормозного нейрона, активированного возбуждающим нейроном, образует синапсы на той клетке, которая ее возбуждает, образуя своеобразную обратную связь тормозного типа. Возвратное торможение обнаружено в двигательных центрах передних рогов спинного мозга и служит, по-видимому, для дозирования возбуждения, приходящего к эфферентным нейронам.
В тех же двигательных центрах спинного мозга обнаружена еще одна форма торможения – реципрокное ((от лат. reciprocus — возвращающийся, обратный, взаимный)) торможение, являющееся постсинаптическим. Такое торможение обеспечивает одновременное возбуждение нейронов мышц-сгибателей и торможение нейронов мышц-разгибателей на той же стороне тела, что необходимо для осуществления движения.
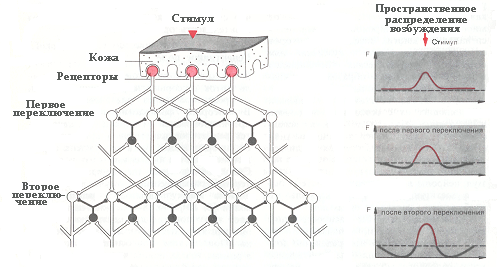
Рис. Эффект латерального торможения. Добавление тормозных вставочных нейронов сужает распределение частоты импульсации на высших уровнях.
Существует и еще одна форма торможения, получившая название латерального торможения, которое может быть как постсинаптическим, так и пресинаптическим. Возбуждающий нейрон через вставочные тормозные клетки вызывает торможение соседних нейронов, что приводит к формированию тормозной зоны, расположенной вокруг возбужденного нейрона. Латеральное торможение играет особо важную роль в сенсорных системах, при выделении наиболее важного признака стимула. Описанные формы торможения представлены на рисунке.
|
|