Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Лекция № 3. 5 страница
|
|
Мочевая кислота является конечным продуктом распада ПУРИНОВЫХ НУКЛ.
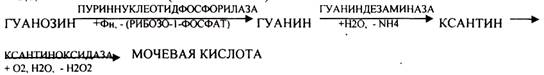
Уровень мочевой кислоты будет свидетельствовать об интенсивности распада ПУРИНОВЫХ оснований тканей организма и пищи. При распаде ПИРИМИДИНОВЫХ оснований из них образуется:
l.B-аланин СО2 NH3 МОЧЕВИНА
 |  |
2.бета -АМИНОИЗОБУТИРАТ.
НАРУШЕНИЕ ОБМЕНА НУКЛЕОТИДОВ.
ГИПЕРУРИКЕМИЯ - повышение уровня мочевой кислоты в крови указывает на повышенный распад нуклеиновых кислот или пуриновых нуклеотидов. Мочевая кислота плохо растворима и выпадает в кристаллы в кислой среде. Развивается подагра, артриты, инициируется развитие МКБ (моче каменной болезни). В этих случаях рекомендуется содовое питьё. Образуются УРАТЫ натрия, которые являются растворимыми и легко выводятся из организма.
ОБРАЗОВАНИЕ КРЕАТИНИНА.
КРЕАТИНИН образуется из креатина, который в свою очередь синтезируется в печени из аминокислот, затем транспортируется в мышечную ткань, где взаимодействует с АТФ.
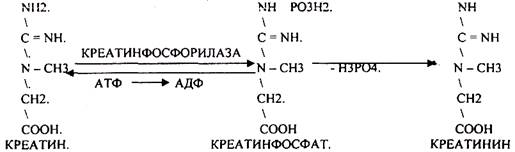
КРЕАТИНИН является конечным продуктом распада креатина. Он поступает в кровь и выделяется с мочой. КРЕАТИНИН не подвергается обратному всасыванию и является показателем фильтрационной способности почек. При её нарушении уровень КРЕАТИНИНА в моче будет понижаться, а в крови - повышаться,._
ОСТАТОЧНЫЙ АЗОТ - это содержание в крови азота всех азотсодержащих соединений, остающихся после осаждения белков. Его содержание колеблется 14 - 28ммоль/л. К остаточному азоту относится:
1.Азот промежуточных продуктов (ПЕПТИДЫ, АО, АК, КРЕАТИН, ИНДОЛ, СКАТОЛ и т.д.)
2.Азот конечных продуктов (МОЧЕВИНА, МОЧЕВАЯ К-ТА, ИНДИКАН, КРЕАТИНИН)—
ГИПЕРАЗОТЕМИЯ - повышение содержания в крови остаточного азота. Она может быть:
1.продукционная - повышение образования компонентов остаточного азота, главным образом
вследствие распада белков.
2.ретенционная - задержка азотистых шлаков из-за нарушения выделительной функции почек.
В клинике чаще встречается РЕТЕНЦИОННАЯ ГИПЕРАЗОТЕМИЯ.
Азотистый баланс - это разность азота, поступившего с пищей, и азота, выделяющегося с мочой. В физиологическом состоянии у здорового человека с пищей поступает 16гр азота и выделяется столько же. Азотистый баланс равен 0. Это называется азотистым равновесием.
57.
Положительный азотистый баланс связан с преобладанием процессов синтеза над процессами распада. Азот задерживается в организме. Характерно для растущего организма или росте опухолей. Отрицательный азотистый баланс указывает на преобладание процессов распада над процессами синтеза. Может наблюдаться в пожилом и старческом возрасте у здоровых людей. В патологии может быть при ожоговой болезни, деструкции опухоли.
ПАТОЛОГИЯ БЕЛКОВОГО ОБМЕНА.
1. Нарушение на этапе поступления белков. В норме в организм должно поступать 80 – 100гр белков. Обязательно поступление всех незаменимых АК.
Клинические проявления при недостатке определённых АК:
Лизин -тошнота, глаукома, повышение чувствительности к шуму: ГИС - понижение гемоглобина; МЕТ - жировая инфильтрация печени и почек.
При общем белковом голодании развивается белковая недостаточность, которая проявляется понижением уровня белка в сыворотке крови, ГИПОПРОТЕИН- и ГИПОГЛОБУЛИНЕМИЕЙ. Понижается онкотическое давление, и вода устремляется в ткани (развиваются отёки).
2. Нарушение на этапе пищеварения:
A) в желудке: ГИПОАЦИДОЗ, следовательно, пепсин неактивен и нарушается распад белков;
АХЛОРГИДРИЯ, следовательно, белки не расщепляются в желудке, подвергаются гниению.
B) в тонком кишечнике:
Связан с поражением ПЖЖ (панкреатиты), следовательно, недостаточность выработки трипсина и других гидролитических ферментов.
C) в толстой кишке:
Запоры, непроходимость, следовательно, усиление процессов гниения.
3. Нарушение МЕЖУТОЧНОГО обмена - нарушение обмена аминокислот в тканях.
A) Приобретенные - часто связаны с отсутствием витаминов (г.о. В6), следовательно, нарушение процессов ПЕРЕАМИНИРОВАНИЯ, ДЕЗАМИНИРОВАНИЯ. Это приводит к ГИПЕРАЦИДОАМИНЕМИИ, АМИНОАЦИДУРИИ.
B) Наследственные. Чаще всего встречаются нарушения обмена ФЕНИЛАЛАНИНА. В норме ФЕН трансформируется в ТИР.
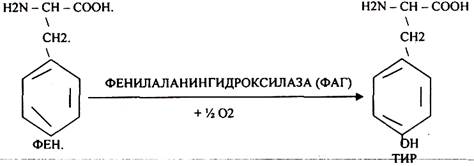
Отсутствие ФАГ приводит к так называемому метаболическому блоку.
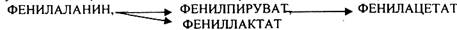
Эти соединения накапливаются в тканях, повышаются в крови, появляются в моче -
ФЕНИЛКЕТОНУРИЯ (ФКУ). Они токсичны, особенно для тканей мозга. Ребёнок отстаёт в умственном развитии ФЕНИЛПИРОВИНОГРАДНАЯ ОЛИГОФРЕНИЯ. Назначается специальная диета, исключающая ФЕНИЛАЛАНИН.
При нарушении метаболизма ТИРОЗИНА не синтезируется МЕЛАНИН, следовательно, нарушается пигментация кожи, радужки. Моча окрашивается в бурый цвет, а затем в чёрный - АЛКАПТОНУРИЯ.
Нарушение на стадии ОРНИТИНОВОГО ЦИКЛА, следовательно накопление промежуточных продуктов (ЦИТРУЛИНА, АРГИНИНСУКЦИНАТА) ЦИТРУЛЛИНУРИЯ, АРГИНИНСУКЦИНАТНОЕ СЛАБОУМИЕ.
Нарушение на уровне выведения конечных продуктов. ЦИСТИНОЗ - нарушение РЕАБСОРБЦИИ АК и повышение их экскреции в 5 -10 раз (ЦИСТИН, ЦИСТЕИН).
58.
Лекция № 19. Матричный биосинтез.
РЕПЛИКАЦИЯ, И ЕЁ ЭТАПЫ.
РЕПАРАЦИЯ ДНК.
ТРАНСКРИПЦИЯ ГЕНА. СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О ГЕНЕ.
СОВРЕМЕННЫЕ МЕТОДЫ ИССЛЕДОВАНИЯ ДНК.
Генетическая догма: информация записана в ДНК и передаётся на дочерние молекулы ДНК
из поколения в поколение с помощью процесса репликации.
ДНК ® РНК ® белок
РЕПЛИКАЦИЯ - процесс самоудвоения ДНК. Этот процесс стал полностью изучен только после того, как УОТСОН и КРИК предложили структуру ДНК в виде двойной спирали, полинуклеотидные цепи которой связаны комплиментарными, азотистыми основаниями (А::: Т, Г::: Ц). Если азотистые основания комплиментарны друг другу, то и полинуклеотидные цепи тоже комплиментарны. В основе механизма репликации лежит принцип комплиментарности. К механизму репликации относится матричный биосинтез. Репликация ДНК идёт полуконсервативным способом: на каждой материнской полинуклеотидной цепи синтезируется дочерняя цепь.
Условия необходимые для репликации:
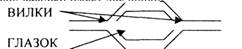
1. Матрица - нити ДНК. Расщепление нити называется РЕПЛИКАТИВНАЯ ВИЛКА. Она
может образовываться внутри молекулы ДНК. Они движутся в разных направлениях,
образуя РЕПЛИКАТИВНЫЙ ГЛАЗОК. Таких глазков в молекуле ДНК ЭУКАРИОТ
несколько, каждый имеет две вилки
2. Субстрат. Пластическим материалом являются ДЕЗОКСИНУКЛЕОТИДТРИФОСФАТЫ:
дАТФ, дГТФ, дЦТФ, дТТФ. Затем происходит их распад до ДЕЗОКСИНУКЛЕОТИДМОНОФОСФАТОВ, двух молекул фосфата неорганического с выделением энергии, т.е. они одновременно являются источником и энергии, и пластического материала.
Д-АТФ® Д-АМФ + ФФ + Е.
3. Ионы магния.
Репликативный комплекс ферментов:
A) ДНК -раскручивающие белки:
1. DNA-A (вызывает расхождение нитей)
2. ХЕЛИКАЗЫ (расщепляют цепь ДНК)
3. ТОПОИЗОМЕРАЗЫ 1 и 2 (раскручивают сверх спирали). Разрывают (3', 5') -
фосфодиэфирные связи. ТОПОИЗОМЕРАЗА 2 у ПРОКАРИОТ называется ГИРАЗА.
B) Белки, препятствующие соединению нитей ДНК (SSB -белки)
C) ДНК-ПОЛИМЕРАЗА (катализирует образование фосфодиэфирных связей). ДНК-
ПОЛИМЕРАЗА только удлиняет уже существующую нить, но не может соединить два свободных НУКЛЕОТИДА.
D) ПРАЙМАЗА (катализирует образование «затравки» к синтезу). Это по своей структуре РНК-ПОЛИМЕРАЗА, которая соединяет одиночные НУКЛЕОТИДЫ.
Е)ДНК-ЛИГАЗА.
5. ПРАЙМЕРЫ - «затравка» для репликации. Это короткий фрагмент, состоящий из РИБОНУКЛЕОТИДТРИФОСФАТОВ (2 - 10). Образование ПРАИМЕРОВ катализируется ПРАЙМАЗОЙ.
Основные этапы репликации.
1.ИНИЦИАЦИЯ репликации.
Происходит под влиянием внешних стимулов (факторов роста). Белки соединяются с рецепторами на плазматической мембране и вызывают репликацию в синтетическую фазу клеточного цикла. Смысл инициации заключается в присоединении в точку репликации DNA-A, стимулирующего расхождение двойной спирали. В этом принимает участие и ХЕЛИКАЗА. Действуют ферменты (ТОПОИЗОМЕРАЗЫ), вызывающие раскручивание сверх спирали. SSB-белки препятствуют соединению дочерних цепей.
Образуется РЕПЛИКАТИВНАЯ ВИЛКА.
Образование дочерних нитей.
Этому предшествует образование ПРАИМЕРОВ с помощью фермента ПРАЙМАЗЫ. Действует ДНК-ПОЛИМЕРАЗА и образуется дочерняя нить ДНК. Этот процесс происходит в соответствии с принципом комплиментарности, и синтез идёт от 5' к 3' концу синтезируемой нити.
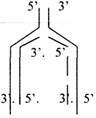
На одной из материнских нитей будет строиться непрерывная цепь, а на противоположной нити - цепь из коротких фрагментов (фрагментов ОКАЗАКИ) Удаление ПРАИМЕРОВ с помощью ЭКЗОНУКЛЕАЗЫ.
Соединение коротких фрагментов с помощью ДНК-ЛИГАЗЫ.
Дочерние нити КОМПЛИМЕНТАРНЫ материнским. Дочерние молекулы ДНК являются копией материнской ДНК. Значение репликации: воспроизведение генотипа в поколениях.
2. РЕПАРАЦИЯ ДНК.
ДНК человека содержит примерно 3 млрд. НУКЛЕОТИДОВ. Если появится одна ошибка на 1млн. НУКЛЕОТИДОВ, то в целом в молекуле ДНК будет 3000 ошибок за один процесс репликации. Ошибки в репликации могут привести к летальному исходу. В организме существует система, исправляющая ошибки - система репарации ДНК. Она состоит из трёх ферментов:
- ЭНДОНУКЛЕАЗА,
- ЭКЗОНУКЛЕАЗА,
- ДНК-ПОЛИМЕРАЗА РЕПАРИРУЮЩАЯ.
- ЭНДОНУКЛЕАЗА вызывает разрыв фосфодиэфирных связей повреждённого НУКЛЕОТИДА с соседним, с той или с другой стороны. В результате образуется два конца. В действие вступает ЭКЗОНУКЛЕАЗА, отщепляющая НУКЛЕОТИДЫ с образующихся концов ДНК в обе стороны. В результате действия этих ферментов на одной нити ДНК возникает дефект. В работу вступает ДНК-ПОЛИМЕРАЗА РЕПАРИРУЮЩАЯ. Она достраивает повреждённую цепь в соответствии с принципом комплиментарности. Т.о. повреждённый НУКЛЕОТИД удаляется.
3. ТРАНСКРИПЦИЯ ГЕНА.
Транскрипция - передача информации с ДНК на РНК (биосинтез РНК). Транскрипции, в отличие от репарации, подвергаются только определённые части молекулы ДНК. Эта часть называется ТРАНСКРИПТОНОМ - фрагмент ДНК, транскрибируемый в РНК. ДНК эукариот прерывистая: участки, несущие информацию (ЭКЗОНЫ), чередуются с участками, не несущими информацию (ИНТРОНЫ). В ДНК с 5'-конца выделяют ПРОМОТОРНУЮ область - место присоединения РНК-ПОЛИМЕРАЗЫ. С 3'-конца - ТЕРМИНАТОРНАЯ зона. Эти области не транскрибируются.
УСЛОВИЯ ТРАНСКРИПЦИИ.
1. Матрица - 1 нить ДНК. Образуется транскрипционный глазок.
2. Структурные компоненты - РИБОНУКЛЕОЗИД-3-ФОСФАТЫ (АТФ, ГТФ, ЦТФ, УТФ). Они будут распадаться до МОНОФОСФАТОВ с выделением энергии.
3. ДНК-зависимая РНК-ПОЛИМЕРАЗА.
ОСНОВНЫЕ ЭТАПЫ ТРАНСКРИПЦИИ.
1. ИНИЦИАЦИЯ.
Заключается в присоединении РНК-ПОЛИМЕРАЗЫ к ПРОМОТОРУ, что приводит к расхождению нитей ДНК. Импульсом к присоединению РНК-ПОЛИМЕРАЗЫ является присоединение ТВР-белка к TATA-САЙТУ.
2. ЭЛОНГАЦИЯ (удлинение).
Соединение РИБОНУКЛЕОЗИДМОНОНУКЛЕОТИДОВ и образование фосфодиэфирных
связей между НУКЛЕОТИДАМИ с помощью РНК-ПОЛИМЕРАЗЫ, которая передвигается
вдоль нити ДНК. Присоединение НУКЛЕТИДОВ идет в соответствии с принципом
комплиментарности, только вместо ДЕЗОКСИНУКЛЕОТИДОВ будут РИБОНУКЛЕОТИДЫ, и вместо ТМФ - УМФ.синтез идёт со скоростью 30 - 50 НУКЛЕОТИДОВ в секунду, пока не дойдёт до Т-зоны.
3. ТЕРМИНАЦИЯ (окончание).Заключается в том, что со стороны 3'-конца образованной РНК присоединяется множество (до 200 - 300) АДЕНИЛОВЫХ НУКЛЕОТИДОВ - поли А. Образуется точная копия гена. АДЕНИЛОВЫЕ НУКЛЕОТИДЫ защищают 3'-конец от действия ЭКЗОНУКЛЕАЗ. С 5'-конца образуется защита, так называемый «САР» (чаще всего УДФ). Эта образовавшаяся копия гена называется ТРАНСКРИПТ.
4. ПРОЦЕССИНГ (созревание).
Заключается в СПЛАЙСИНГЕ удаление неинформативных участков, и соединение ЭКЗОНОВ между собой. Играет важную роль в эволюции организмов, т.к. возможен альтернативный СПЛАЙСИНГ. Это свидетельствует о том, что из одной пре-ИРНК образуется несколько ИРНК и соответственно несколько белков, что проявляется в разнообразии признаков у организмов.
Лекция № 20. Биосинтез белка.
1.БЕЛОКСИНТЕЗИРУЮЩИЙ АППАРАТ.
2.ТРАНСЛЯЦИЯ.
3.РЕГУЛЯЦИЯ БИОСИНТЕЗА БЕЛКА.БИОСИНТЕЗ БЕЛКА.
Трансляция или собственно биосинтез белка - это перевод генетического текста М-РНК в последовательность аминокислот в белке. Характеристика белоксинтезирующего аппарата клетки.
1. МРНК - источник информации.
У эукариот имеется особенность - САР (шапка, кепка), представленная МЕТИЛ-ГТФ. САР защищает МРНК от гидролиза и способствует её соединению с РИБОСОМАМИ. С САР связываются САР-связывающие белки. На МРНК находится стартовый кодон, представленный триплетом АУГ, представленный аминокислотой - МЕТ.
РИБОСОМЫ - комплексы РРНК с порядка 80 белками, включая ферменты.
РИБОСОМА состоит из двух единиц- большой и малой. Она у ЭУКАРИОТ более крупная,
её формула 80S (40S и 60S). У ПРОКАРИОТ она имеет формулу 70S (30S и 50S)
Аминокислоты (20 видов).
ТРНК(31 вид).
Несоответствие числа транспортных РНК и числа кодонов (61) снимается за счёт возможности узнавания одной транспортной РНК нескольких кодонов данной аминокислоты (ГЛИЦИН имеет три варианта ДНК-кода - ГГУ, ГГЦ, ГГА). Все эти 3 варианта узнаются одним видом Т-РНК - ЦЦН. В состав транспортной РНК входят минорные азотистые основания, способные узнавать вариабельный участок разных кодонов одной и той же аминокислоты. Транспортная РНК выполняет функцию АДАПТОРА между МРНК и белком.
Фермент синтеза комплекса транспортной РНК с аминокислотой - АМИНОАЦИЛ-ТРНК-СИНТЕТАЗА (требует энергии АТФ).
Белковые факторы:
факторы ИНИЦИАЦИИ (ФИ) начала трансляции, факторы ЭЛОНГАЦИИ (ФЭ) - продолжатели, факторы ВЫСВОБОЖДЕНИЯ (R-факторы).
Ионы магния, как КОФАКТОРЫ.
АТФ, ГТФ - поддержка энергией.
Этапы трансляции:
1.РЕКОГНИЦИЯ (распознавание) - узнавание между аминокислотами и их транспортной
РНК.
2.АК + ТРНК аминоацил-т-РНК-синтетаза АК-ТРНК
 АТФ®АМФ + ФФ МЕТИОНИЛ-ТРНК
АТФ®АМФ + ФФ МЕТИОНИЛ-ТРНК
РЕКОГНИЦИЯ происходит столько раз, сколько аминокислот входит в состав белка.
3. ИНИЦИАЦИЯ - начало процесса трансляции.
На этом этапе РИБОСОМА взаимодействует с МРНК и находит стартовый кодон. Малая её единица взаимодействует с МЕТИОНИЛ-ТРНК и образует инициирующий комплекс, способный распознавать стартовый кодон. Этому предшествует разделение РИБОСОМЫ с помощью ФИ-3. Образование инициирующего комплекса происходит с помощью ФИ-2. Затем инициирующий комплекс присоединяется к МРНК с 5'-конца. Узнаванию 5'-конца способствуют САР и САР-связывающие белки. Реакцию обеспечивают ФИ-1. Сканирование инициирующим комплексом матричной РНК путём продвижения от 5' к 3'-концу до обнаружения стартового кодона антикодоном МЕТ-ТРНК. Данный процесс энергозависим, требует энергии АТФ. После обнаружения стартового кодона собирается полностью РИБОСОМА путём фиксации 60S единицы, и высвобождаются белковые ФИ-1, 2, 3 и САР-связывающие белки. В РИБОСОМЕ выделяют Р-участок и А-участок. Р-участок (ПЕПТИДИЛЬНЫЙ) - в нём происходит образование ПЕПТИДНЫХ связей. Это закрытая область РИБОСОМЫ. Вход в неё извне запрещён. А-участок (АМИНОАЦИЛЬНЫЙ). Это открытая область РИБОСОМЫ - для поступления следующей аминокислоты.
4. ЭЛОНГАЦИЯ (продолжение) протекает циклически в виде последовательной смены трёх
фаз:
- присоединение следующей АМИНОАЦИЛ-ТРНК в соответствии со смыслом следующего кодона. Для процесса требуется энергия ГТФ и ФЭ-1 (проникновение в РИБОСОМУ).
- ПЕПТИЗАЦИЯ. Фермент ПЕПТИДИЛТРАНСФЕРАЗА образует ПЕПТИДНУЮ связь между двумя аминокислотными остатками и одновременно разрушает сложноэфирную связь между первой аминокислотой и её ТРНК. В результате идёт образование растущего ПЕПТИДА в А-участке и высвобождение первой ТРНК.
- ТРАНСЛОКАЦИЯ (перемещение).
При этом происходит перемещение РИБОСОМЫ на один кодон в направлении 3'-конца. При этом все остальные компоненты (МРНК, ТРНК) остаются на месте. Для процесса требуется энергия ГТФ и белковый ФЭ-2. Процесс циклический, т.е. фазы чередуют друг друга. Это происходит до обнаружения стоп (нонсенс)-кодона. Он не кодирует ни одну аминокислоту. Элонгация становится невозможной. Элонгацию и инициацию обозначают как собственно трансляцию.
5. ТЕРМИНАЦИЯ (прекращение).
Стоп -кодон распознаётся R-факторами (РЕЛИЗИНГ). Эти факторы высвобождают из связи все компоненты белоксинтезирующего комплекса: РИБОСОМУ, МРНК, ПОЛИПЕПТИД. Помогает фермент - ПЕПТИДИЛТРАНСФЕРАЗА, отщепляющий транспортную РНК от образованного ПОЛИПЕПТИДА. Для ТЕРМИНАЦИИ нужна энергия ПГФ. РИБОСОМА может повторно использоваться в трансляции. Матричная РНК или повторно используется в трансляции, или гидролизуется. ПОЛИПЕПТИД вступает в этап ПРОЦЕССИНГА белка.
6. БРОЦЕССИНГ белка (созревание) совокупность химических модификаций
ПОЛИПЕПТИДА, заканчивающихся формированием зрелой белковой молекулы. ПРОЦЕССИНГ белка может быть:
КОНТРАНСЛЯЦИОННЫЙ (во время трансляции)
ПОСТТРАНСЛЯЦИОННЫЙ.
Варианты ПРОЦЕССИНГА:
A) ограниченный протеолиз: отщепление N-концевой аминокислоты (МЕТ), отщепление ПЕПТИДНОГО фрагмента.
АЦИЛИРОВАНИЕ (присоединение остатка СНЗСООН)
ФОСФОРИЛИРОВАНИЕ
ГЛИКОЗИЛИРОВАНИЕ - образование ГЛИКОПРОТЕИНОВ и ПРОТЕОГЛИКАНОВ.
E) ГИДРОКСИЛИРОВАНИЕ аминокислот
F) ОКИСЛЕНИЕ аминокислот
G) Образование четвертичной структуры в случае ОЛИГОМЕРНОСТИ белка.
ПРОЦЕССИНГ включает в себя наивысшую точку - ФОЛДИНГ - сворачивание, обретение
белком высших уровней пространственно-структурной организации, заканчивающееся
формированием структурно и функционально зрелой молекулы.
ШАПЕРОНЫ - над молекулярные комплексы белковой природы, способствующие быстрому и
правильному ФОЛДИНГУ. В большом числе представлены белками теплового шока.
ШАПЕРОНЫ препятствуют приобретению белком неправильной конформации. Неудачный ФОЛДИНГ заканчивается появлением аномальных белков, которые должны быть элиминированы.
УБИКВИТИН белок, присутствующий в каждой клетке организма. Это «билет» на уничтожение аномального белка. Белок, меченый УБИКВИТИНОМ, разрушается в ПРОТЕОСОМАХ. В норме обеспечивается гомеостаз структуры белка, сохранение его нативной структуры. При заболеваниях образуются аномальные белки, которые не утилизируются - ПРИОНЫ -ПРОТЕИНОГЕННЫЕ, информативные частицы. Это продукты неправильного ФОЛДИНГА, которые, попадая в организм извне, трансформируют нормальные белки данного организма в виде цепной реакции. ПРИОННЫЕ белки не расщепляются в ЖКТ, а всасываются в неизменённом виде. По сути ПРИОНЫ - это отрицательные ШАПЕРОНЫ. КОМПАРТМЕНТАЛИЗАЦИЯ БЕЛКОВ (механизмы адресования).
Белки должны быть правильно распределены в клетке или выделены из неё на экспорт. За этот процесс отвечает сигнальный участок - это фрагмент аминокислотной последовательности синтезированного ПОЛИПЕПТИДА, содержащего условный адрес размещения белка. Известно, что преобладание гидрофобных аминокислот в сигнальном участке, направляет белок в мембранную структуру клетки. Преобладание гидрофильных аминокислот в сигнальном участке способствует проникновению белка в цитоплазму и выделение на
ПРЕПРОИНСУЛИН - ПОЛИПЕПТИД, содержащий 110 аминокислотных остатков. Он содержит сигнальный участок, представленный гидрофильными аминокислотами, которые адресуют ПРЕПРОИНСУЛИН в ЭПР. После этого сигнальный участок отщепляется и образуется ПРОИНСУЛИН, содержащий 84 аминокислотных остатка. Он подвергается ОГРАНИЧЕННОМУ ПРОТЕОЛИЗУ путём вырезки внутреннего участка С-ПЕПТИДА. Образуется две цепочки из 21 и 30 аминокислот, которые затем соединяются дисульфидными связями с образованием молекулы инсулина из 51 аминокислотного остатка.
РЕГУЛЯЦИЯ БИОСИНТЕЗА БЕЛКА.
Несмотря на единство общего генотипа, в организме человека присутствует примерно 200 фенотипов клеток, и фенотипические различия определяются экспрессией генов. В каждой клетке, независимо от фенотипа, экспрессируются гены «домашнего хозяйства» обеспечивающие элементарные процессы жизнедеятельности, характерные для каждой клетки. Регуляция биосинтеза белка у ПРО- и ЭУКАРИОТ различна. У ПРОКАРИОТ основные положения теории регуляции изложены в 1961г. Ф. ЖАКОБОМ и Ж. МОНО.
1. Регуляция происходит только на уровне транскрипции. Первичные транскрипты генов у них
транслируются до завершения транскрипции.
2. Неоднородность ГЕНОМОВ. В геноме есть структурные гены и есть регуляторные области,
которые могут включать регуляторные элементы и регуляторные гены. Структурные гены
кодируют синтез структурных и функциональных белков. Регуляторные элементы не
кодируют синтез белков вообще, но влияют на процесс транскрипции.
Регуляторными элементами являются:
- ПРОМОТОР - место прикрепления к ДНК РНК-ПОЛИМЕР АЗЫ, ОПЕРАТОР - место взаимодействия регуляторных белков с ДНК. Регуляторные гены кодируют синтез регуляторных белков. К ним относится белок -РЕПРЕССОР, который Может блокировать считывание информации, связываясь с оператором. Фрагмент ДНК, подверженный транскрипции называется ОПЕРОН (ПРОМОТОР, ОПЕРАТОР, структурный ген). За пределами ОПЕРОНА находятся гены-регуляторы, кодирующие синтез белка - РЕПРЕССОРА.
3. Регуляция биосинтеза белков у ПРОКАРИОТ протекает альтернативно путём репрессии и индукции.
ПРИМЕР: ЛАКТОЗНЫЙ ОПЕРОН. В микробной клетке лактоза с помощью лактазы расщепляется до галактозы и глюкозы. Лактозный ОПЕРОН регулирует синтез лактазы. Если в среде присутствует лактоза, то БЕЛОК-РЕПРЕССОР вытесняется из связи с оператором и гены лактазы транскрибируются. Лактоза выступает индуктором.
Регуляция биосинтеза белка у ЭУКАРИОТ происходит на всех уровнях матричных биосинтезов.
1. На уровне транскрипции - групповая репрессия гистонами. У человека 90% ДНК репрессировано.
2. Амплификация генов - повышение числа копий гена в геноме (повышается площадь транскрипции).
3. Регуляция транскрипции сигналами-регуляторами (усилителями и душителями). Для сигналов усилителей принят термин ЭНХАЙСЕР. Они не кодируют синтез белка, чрезвычайно эффективны, может наблюдаться 200-кратное усиление транскрипции. Действие не специфично (одновременно может влиять на группу генов). Значительно удалён от ПРОМОТОРА гена-мишени. Подвержен влиянию регуляторных факторов (гормонов). Сигналы-душители называются САЙЛЕНСЕРЫ - угнетатели транскрипции. Особенности действия подобны ЭНХАЙСЕРАМ, только действие противоположное.
4. Регуляция на уровне процессинга МРНК разрешение или запрещение процессинга
дифференциальный процессинг включает альтернативный СПЛАЙСИНГ - сборка РНК из разных экзонов, и редактирование МРНК - замена одного из нуклеотидов с изменением генетической информации, приводящее к образованию изменённых белков
5. На уровне стабильности и активности МРНК. МРНК в клетке образует комплекс с белками, который называется ИНФОРМОСОМА. В их составе МРНК не разрушается ферментами, сохранения в активном, стабильном состоянии. При необходимости она высвобождается из комплекса и транслируется. Процесс образования и распада ИНФОРМОСОМ регулируется гормонами. С одной молекулы РНК транслируется большое количество белков.
6. Регуляция на уровне трансляции:
Тотальная регуляция может быть в виде тотальной репрессии или индукции за счёт
изменения концентрации белковых факторов трансляции
Избирательная дискриминация. Определённые виды МРНК избирательно не
транслируются. С них не синтезируется белок. Трансляция с альтернативных стартовых участков. Т.о. образом может происходить выбор исходной стартовой точки трансляции.
Лекция №21.
Ключевые позиции и проблемы молекулярной биологии геномного периода.
1. ПЕРВЫЕ ПРЕДСТАВЛЕНИЯ О ГЕНЕ.
2. СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О ПРИРОДЕ БИОХИМИЧЕСКОГО ПОЛИМОРФИЗМА.
3. СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ СТРУКТУРНО-ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ ДНК.
Прочтение генома привело к формированию новой биологии. Предпосылки возможности биологии нового времени: 1953г. Д. УОТСОН и Ф. КРИК открыли структуру ДНК. 1961г. Ф. КРИК расшифровал генетический код. 1970г. Г. ТЕМЕН и Д. БАЛТИМОР открыли ОБРАТНУЮ ТРАНСКРИПТАЗУ (способность на основе РНК синтезировать ДНК). Значение: возможность изучения изолированных генов. 1983г. К. МЮЛЛИС - реакция PCR (ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ) - управляемое тиражирование ДНК в пробирке. 2001г. первая версия генома человека.
ЭТАПЫ И ПРИНЦИПЫ PCR.
PCR - это метод, позволяющий в пробирке получать любое количество копий заданного участка молекулы ДНК. Стадии метода PCR:
1. Выделение ДНК из биологического материала.
2. Амплификация - репликация на органическом участке молекулы ДНК. Производится за счёт
работы ферментов и смены температурных режимов.
3. ДЕТЕКЦИЯ продуктов PCR (копий заданного участка)
Схема PCR:
ДНК
Т= 90, Денатурация (расплетение) ДНК
Т=50. Отжиг ПРАЙМЕРОВ
Т=70. Синтез ДНК
ПРАЙМЕРЫ специфические ОЛИГОНУКЛЕОТИДНЫЕ последовательности комплиментарные изучаемому участку ДНК. Они синтезируются в искусственных условиях на основе НУКЛЕОТИДНОЙ последовательности ДНК. ПРАЙМЕРЫ ограничивают зону копирования, являются закладками в ДНК. Синтез ДНК идёт с помощью ДНК-ПОЛИМЕРАЗЫ. Этот фермент выделен из микроорганизмов, обитающих в горячих источниках. Т.о. в результате первого цикла вместо одной молекулы ДНК образуется две. На следующий цикл матрицей будут являться все продукты предшествующего цикла. За 20 - 30 циклов количество фрагментов ДНК вырастает в 1000000 раз. Эти реакции производят в АМПЛИФИКАТОРЕ - приборе, где чередуются циклы нагревания и охлаждения. Продолжительность каждого этапа измеряется секундами. Факторы необходимые для PCR:
1. изучаемая ДНК,
2. ПРАЙМЕРЫ изучаемого участка,
3.Термостабильная ДНК-ПОЛИМЕРАЗА,
4. Строительный материал. Он же является источником энергии,
5. Оборудование для АМПЛИФИКАЦИИ.
Сферы применения PCR.
В 1993г. К. МЮЛЛЕСУ была присуждена Нобелевская премия в области химии, за открытие
PCR.
1. изучение генома микроорганизмов
2. клиническая лабораторная диагностика. PCR даёт возможность обнаружения
наследственных заболеваний, а также обнаружить чужеродный геном в тканях человека
(микроорганизмы, вирусы и т.д.). Реакция пойдёт или не пойдёт. В настоящее время PCR - самый точный метод микробиологической диагностики.
СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О СТРУКТУРНО-ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ ДНК. 95% ДНК человека представляет не генная часть. 5% - собственно гены. Не генная часть представлена:
1. ТАНДЕМНЫЕ ПОВТОРЫ монотонные повторы НУКЛЕОТИДОВ, как правило дуплетного характера, не имеющие смысла. Это так называемые «пустынные участки» ДНК. В настоящее время смысл этих участков: выполнение структурной функции и площадки для образования генов в эволюции (эволюционный резерв).
2. ПСЕВДОГЕНЫ - неактивные, но стабильные генетические элементы, возникающие в результате мутации в ранее работавших генах (гены, выключенные мутацией). Это побочный
продукт и генетический резерв эволюции. Составляют 20 - 30% не генной части ДНК.
3. Мобильные генетические элементы:
-ТРАНСПОЗОНЫ - участки ДНК, способные вырезаться и встраиваться в другие области
ДНК. Это так называемые «странники генов».
-РЕТРОТРАНСПОЗОНЫ - участки ДНК, копирующиеся в пределах генома, как внутри
хромосомы, так и между ними. Могут изменять смысл структурных генов человека, приводят к мутациям. Геном человека изменяется в течении жизни на 10 - 30%.
- поврежденные неактивные, мобильные генетические элементы. Не могут ни вырезаться, ни встраиваться из-за отсутствия в клетке ОБРАТНОЙ ТРАНСФЕРАЗЫ. Если фрагмент поступает в клетку с вирусом, то тогда эти гены начинают транскрибироваться. ФУНКЦИОНАЛЬНЫЕ ЭЛЕМЕНТЫ ГЕНОМА:
1. СТРУКТУРНЫЕ ГЕНЫ
2. РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ
Структурные гены кодируют синтез МРНК, ТРНК, РРНК.
Регуляторные элементы не кодируют РНК и, соответственно, белков; влияют на работу
структурных генов.
ГЕНОМИКА - отрасль молекулярной биологии, изучающая структуру и механизмы работы гена. В настоящее время установлено и изучено 35000 генов человека. Из них в каждой клетке работает 25%.
СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О ПРИРОДЕ БИОХИМИЧЕСКОГО ПОЛИМОРФИЗМА.
В 40-е годы 20в БИДЛОМ и ТЕЙТУМОМ выдвинут принцип: 1 ген - I фермент. Они исходили из общей схемы биосинтеза белка, однако этот принцип не объясняет:
1. Фенотипическое различие между клетками,
2. Индивидуальные различия между организмами одного вида. Каждый человек отличается от другого человека на 0, 1% генома.
|
|