Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Тема 4: Физиология эндокринной системы
|
|
Физиология гипоталамо-гипофизарно-надпочечниковой системы
Время: 2 часа.
Учебная цель: уяснить особенности гуморальной регуляции, свойства и механизм действия гормонов, саморегуляторный принцип выделения гормонов. Усвоить значение гипоталамо-гипофизарно-надпочечниковой системы в адаптивных реакциях и в регуляции различных функций организма.
Содержание занятия
1. Железами внутренней секреции, или эндокринными железами, являются специализированные органы, которые выделяют образующиеся в них продукты секреции непосредственно в кровь или тканевую жидкость. Это: гипофиз, щитовидная железу, околощитовидные железы, корковое и мозговое вещество надпочечников, островковый аппарат поджелудочной железы, половые железы, тимус, и эпифиз. Эндокринной активностью обладает также плацента. Кроме того, эндокринные клетки могут присутствовать в пищеварительном тракте, почках, сердечной мышце, вегетативных ганглиях. Эти клетки образуют так называемую диффузную эндокринную систему. Общей функцией для всех желез внутренней секреции является выработка гормонов.
Гормоны (греч. Ο ρ μ ό ν η) (греч. hormao — возбуждаю, побуждаю) — биологически активные химические вещества, выделяемые эндокринными железами непосредственно в организме и оказывающие значительные физиологические эффекты на организм в целом либо на определённые органы и ткани-мишени. Гормоны служат гуморальными (переносимыми с кровью) регуляторами определённых процессов в различных органах и системах.
Химическая природа гормонов и биологически активных веществ различна. От сложности строения гормона зависит продолжительность его биологического действия, например, от долей секунды у медиаторов и пептидов до часов и суток у стероидных гормонов и йодтиронинов.
Классификация гормонов и биологически активных веществ (БАВ) по химической структуре:
- Производные аминокислот:
производные тирозина: тироксин, трийодтиронин, дофамин, адреналин, норадреналин;
производные триптофана: мелатонин, серотонин;
производные гистидина: гистамин. - Белково-пептидные гормоны:
полипептиды: глюкагон, кортикотропин, меланотропин, вазопрессин, окситоцин, пептидные гормоны желудка и кишечника;
простые белки (протеины): инсулин, соматотропин, пролактин, паратгормон, кальцитонин;
сложные белки (гликопротеиды): тиреотропин, фоллитропин, лютропин. - Стероидные гормоны:
кортикостероиды (альдостерон, кортизол, кортикостерон);
половые гормоны: андрогены (тестостерон), эстрогены и прогестерон. - Производные жирных кислот:
арахидоновая кислота и ее производные: простагландинм: простациклины, тромбоксаны, лейкотриены.
Функциональная классификация гормонов:
- Эффекторные гормоны — гормоны, которые оказывают влияние непосредственно на орган-мишень.
- Тропные гормоны — гормоны, основной функцией которых является регуляция синтеза и выделения эффекторных гормонов. Выделяются аденогипофизом.
- Рилизинг-гормоны — гормоны, регулирующие синтез и выделение гормонов аденогипофиза, преимущественно тропных. Выделяются нервными клетками гипоталамуса.
Несмотря на то, что гормоны имеют разное химическое строение, для них характерны некоторые общие биологические свойства.
Общие свойства гормонов:
- Строгая специфичность физиологического действия.
- Высокая биологическая активность: гормоны оказывают свое физиологическое действие в чрезвычайно малых дозах.
- Дистантный характер действия: клетки-мишени располагаются обычно далеко от места образования гормона.
- Многие гормоны (стероидные и производные аминокислот) не имеют видовой специфичности.
- Генерализованность действия.
- Пролонгированность действия.
Различают следующие варианты действия гормонов:
- гормональное, или гемокринное, т.е. действие на значительном удалении от места образования;
- изокринное, или местное, когда химическое вещество, синтезированное в одной клетке, оказывает действие на клетку, расположенную в тесном контакте с первой, и высвобождение этого вещества осуществляется в межтканевую жидкость и кровь;
- нейрокринное, или нейроэндокринное (синаптическое и несинаптическое), действие, когда гормон, высвобождаясь из нервных окончаний, выполняет функцию нейротрансмиттера или нейромодулятора, т.е. вещества, изменяющего (обычно усиливающего) действие нейротрансмиттера;
- паракринное - разновидность изокринного действия, но при этом гормон, образующийся в одной клетке, поступает в межклеточную жидкость и влияет на ряд клеток, расположенных в непосредственной близости;
- юкстакринное – разновидность паракринного действия, когда гормон не попадает в межклеточную жидкость, а сигнал передается через плазматическую мембрану рядом расположенной другой клетки;
- аутокринное действие, когда высвобождающийся из клетки гормон оказывает влияние на ту же клетку, изменяя ее функциональную активность;
- солинокринное действие, когда гормон из одной клетки поступает в просвет протока и достигает таким образом другой клетки, оказывая на нее специфическое воздействие (например, некоторые желудочно-кишечные гормоны).
Каждый гормон не работает в одиночку. Поэтому необходимо учитывать возможные результаты их взаимодействия.
Синергизм — однонаправленное действие двух или нескольких гормонов. Например, адреналин и глюкагон активируют распад гликогена печени до глюкозы и вызывают увеличение уровня сахара в крови.
Антагонизм всегда относителен. Например, инсулин и адреналин оказывают противоположные действия на уровень глюкозы в крови. Инсулин вызывает гипогликемию, адреналин — гипергликемию. Биологическое же значение этих эффектов сводится к одному — улучшению углеводного питания тканей.
Пермиссивное действие гормонов заключается в том, что гормон, сам не вызывая физиологического эффекта, создает условия для ответной реакции клетки или органа на действие другого гормона. Например, глюкокортикоиды, не влияя на тонус мускулатуры сосудов и распад гликогена печени, создают условия, при которых даже небольшие концентрации адреналина увеличивают артериальное давление и вызывают гипергликемию в результате гликогенолиза в печени.
2. Когда гормон, находящийся в крови, достигает клетки-мишени, он вступает во взаимодействие со специфическими рецепторами. Каждому конкретному гормону соответствуют исключительно «свои» рецепторы, находящиеся в конкретных органах и тканях — только при взаимодействии гормона с ними образуется гормон-рецепторный комплекс.
Существуют два основных механизма действия гормонов на уровне клетки:
- Реализация эффекта с наружной поверхности клеточной мембраны.
- Реализация эффекта после проникновения гормона внутрь клетки.
Первый тип гормонов соединяется с рецепторами, находящимися внутри клеток — как правило, в цитоплазме. К ним относятся гормоны с липофильными свойствами — например, стероидные гормоны (половые, глюко- и минералокортикоиды), а также гормоны щитовидной железы. Будучи жирорастворимыми, эти гормоны легко проникают через клеточную мембрану и начинают взаимодействовать с рецепторами в цитоплазме или ядре. В этой группе гормонов гормон-рецепторный комплекс, образовавшись в клетке, начинает взаимодействовать с хроматином ядра и тем самым ускоряет или замедляет работу тех или иных генов и корректирует процессы метаболизма.
Второй тип гормонов характеризуются тремя особенностями:
- они растворяются в воде;
- не связываются с белками-носителями;
- начинают гормональный процесс, как только соединяются с рецептором, который может находиться в ядре клетки, ее цитоплазме или располагаться на поверхности плазматической мембраны.
В механизме действия гормон-рецепторного комплекса таких гормонов обязательно участвуют посредники, которые индуцируют ответ клетки. Наиболее важные из таких посредников — цАМФ (циклический аденозинмонофосфат), инозитолтрифосфат, ионы кальция. Участие ионов кальция как посредника обеспечивает воздействие на клетки таких гормонов, как вазопрессин и катехоламины.
Выполнив свою задачу, гормоны либо расщепляются в клетках-мишенях или в крови, либо транспортируются в печень, где расщепляются, либо, удаляются из организма в основном с мочой (например, адреналин).
3. Для поддержания соответствующей концентрации гормона, эндокринный орган постоянно должен получать " информацию" о системной концентрации гормона или какой либо его функции. Эта " информация" поступает в эндокринную систему благодаря существованию механизма обратной связи, которая обеспечивает передачу сведений о периферической концентрации гормона в крови обратно в секретирующий орган.
Система отрицательных обратных связей принимает участие в регуляции секреции буквально всех эндокринных органов. Положительная обратная связь проявляется значительно реже, но она также имеет место при определенных эндокринных состояниях.
Каждый тропный гормон передней доли гипофиза находится под отрицательным контролем гормонов, которые секретируются их органами-мишенями. Контролируемой переменной является концентрация гормонов в плазме, секретируемых органом-мишенью (надпочечники, гонады или щитовидная железа).
Скорость секреции гормона регулируется стимулирующим действием тропного гормона, вырабатываемого передней долей гипофиза, которая, в свою очередь, регулируется рилизинг-гормоном, секретируемым нейронами гипоталамуса. Этот отдел мозга содержит:
1) нейроны, способные реагировать на изменение концентрации в крови гормонов из органов-мишеней;
2) нулевую точку отсчета, соответствующую нормальной концентрации этих гормонов в крови.
Гипоталамус сравнивает эти два значения и, если концентрация циркулирующего гормона из органа-мишени меньше нулевой точки отсчета (нормальной концентрации), он увеличивает секрецию соответствующего рилизинг-гормона. Это повышает секрецию тропного гормона, который стимулирует секреторную активность органа-мишени до тех пор, пока концентрация гормона в плазме, вырабатываемого этим органом, не вернется к нормальному уровню. Аналогично, если концентрация гормона в плазме превышает некий определенный уровень, каскад реакций претерпевает обратные изменения, и концентрация гормона падает. При этом гормоны из органов-мишеней обнаруживают отрицательную обратную связь, и их секреция подавляется в результате снижения секреции тропных или рилизинг-гормонов или снижения секреции тех и других.
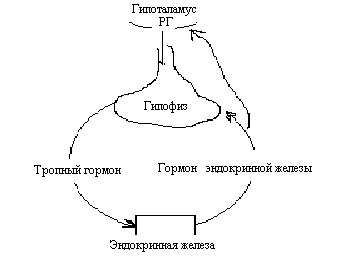
4. Гипоталамус - участок промежуточного мозга расположенный ниже таламуса служит местом непосредственного взаимодействия высших отделов ЦНС и эндокринной системы. В нервных клетках гипоталамуса образуются вещества, которые по системе портальных капилляров достигают гипофиза и регулируют секрецию гипофизарных гормонов, точнее их освобождение. Эти вещества получили сначала наименование нейрогормонов, а затем рилизинг-факторов (от англ.release – освобождать), или либеринов. Вещества с противоположным действием, т.е. угнетающие освобождение гипофизарных гормонов, стали называть ингибирующими факторами, или статинами.
К настоящему времени в гипоталамусе открыто 7 стимуляторов и 3 ингибитора секреции гормонов гипофиза, а именно: кортиколиберин, тиролиберин, люлиберин, фоллилиберин, соматолиберин, пролактолиберин, меланолиберин, соматостатин, пролактостатин и меланостатин.
В супраоптическом и паравентрикулярном ядрах гипоталамуса секретируются гормоны окситоцин и вазопрессин. Нейроны, составляющие эти ядра, имеют длинные аксоны, которые в составе ножки гипофиза образуют гипоталамо-гипофизарный тракт и достигают задней доли гипофиза. Синтезированные в гипоталамусе окситоцин и вазопрессин доставляются в нейрогипофиз путем аксонального транспорта с помощью специального белка-переносчика, получившего название «нейрофизин».
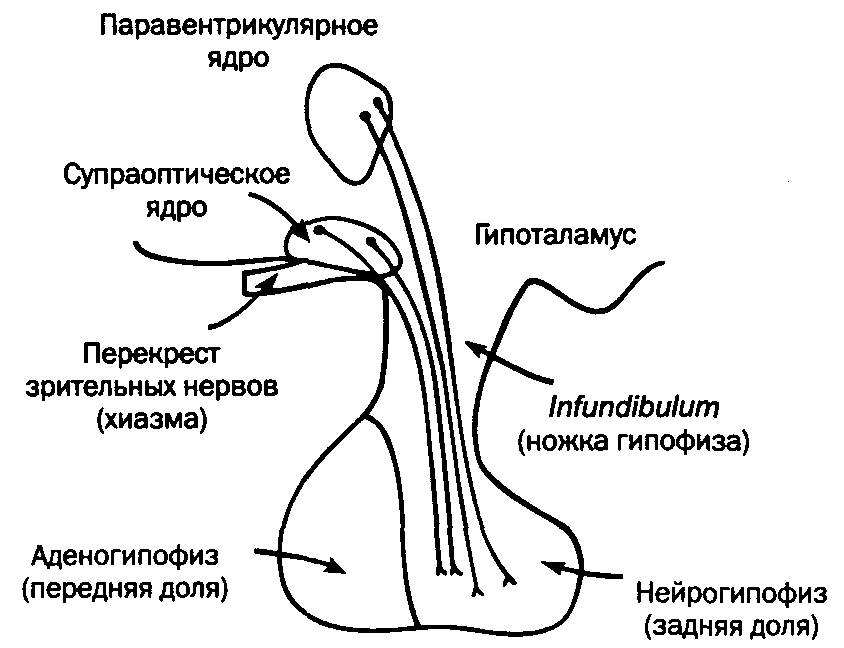
Гипофиз, hypophysis (glandula pituitaria), — небольшая железа, округлой формы, связанная с головным мозгом, посредством гипофизарной ножки. Железа расположенна в турецком седле, где укреплена посредством diaphragma sellae turcicae. Гипофиз также называют нижним придатком мозга. В придатке мозга различают 2 доли: переднюю, lobus anterior (adenohypophysis), и заднюю, lobus posterior (neurohypophysis). Гипофиз регулирует активность ряда желез внутренней секреции и служит местом выделения гипоталамических гормонов крупноклеточных ядер гипоталамуса. Состоит из двух эмбриологически, структурно и функционально различных частей - нейрогипофиза - выроста промежуточного мозга и аденогипофиза, ведущей тканью которого служит эпителий.
5, 6. В аденогипофизе вырабатывается 6 гормонов, из них 4 являются тропными (адренокортикотропный гормон, или кортикотропин, тиреотропный гормон, или тиреотропин и 2 гонадотропина — фолликулостимулирующий и лютеинизирующий гормоны), а 2 — эффекторными (соматотропный гормон, или соматотропин, и пролактин).
Адренокортикотропный гормон (АКТГ), или кортикотропин, оказывает стимулирующее действие на кору надпочечников. В большей степени его влияние выражено на пучковую зону, что приводит к увеличению образования глюкокортикоидов, в меньшей — на клубочковую и сетчатую зоны, поэтому на продукцию минералокортикоидов и половых гормонов он не оказывает значительного воздействия.
Тиреотропный гормон (ТТГ), или тиреотропин, активирует функцию щитовидной железы, вызывает гиперплазию ее железистой ткани, стимулирует выработку тироксина и трийодтиронина. Образование тиреотропина стимулируется тиреолиберином гипоталамуса, а угнетается соматостатином. Секреция тиреолиберина и тиреотропина регулируется йодсодержащими гормонами щитовидной железы по механизму обратной связи.
Фолликулостимулирующий гормон (ФСГ), или фоллитропин, вызывает рост и созревание фолликулов яичников и их подготовку к овуляции. У мужчин под влиянием ФСГ происходит образование сперматозоидов. Лютеинизирующий гормон (ЛГ), или лютропин, способствует разрыву оболочки созревшего фолликула, т.е. овуляции и образованию желтого тела, стимулирует выработку прогестерона у женщин и тестостерона у мужчин.
Соматотропный гормон (СТГ), или соматотропин, или гормон роста, принимает участие в регуляции процессов роста и физического развития. Стимуляция процессов роста обусловлена способностью соматотропина усиливать образование белка в организме, повышать синтез РНК, усиливать транспорт аминокислот из крови в клетки. Соматотропин влияет на углеводный обмен, оказывая инсулиноподобное действие. Гормон усиливает мобилизацию жира из депо и использование его в энергетическом обмене.
Пролактин стимулирует рост молочных желез и способствует образованию молока. Гормон стимулирует синтез белка — лактальбумина, жиров и углеводов молока. Пролактин стимулирует также образование желтого тела и выработку им прогестерона. Влияет на водно-солевой обмен организма, задерживая воду и натрий в организме, усиливает эффекты альдостерона и вазопрессина, повышает образование жира из углеводов.
В нейрогипофизе происходит депонирование окситоцина и антидиуретического гормона (вазопрессин).
Антидиуретический гормон (АДГ), или вазопрессин, осуществляет в организме 2 основные функции. Первая функция заключается в его антидиуретическом действии, которое выражается в стимуляции реабсорбции воды в дистальном отделе нефрона. Недостаточная секреция АДГ приводит к развитию несахарного мочеизнурения (diabetes insipidus), основными проявлениями которого являются сильная жажда (полидипсия) и потеря большого количества жидкости с выделяемой мочой (полиурия). В больших дозах (фармакологических) АДГ суживает артериолы, в результате чего повышается артериальное давление.
Окситоцин оказывает стимулирующее действие на гладкую мускулатуру матки и на лактирующую молочную железу. Он усиливает сокращение миоэпителиальных клеток в молочных железах и тем самым способствует выделению молока. Окситоцин является гормоном, обеспечивающим нормальное протекание родового акта (отсюда произошло и его название — от лат. oxy — сильный, tokos — роды). Адекватное проявление этого эффекта возможно при условии достаточной концентрации в крови эстрогенов, которые усиливают чувствительность матки к окситоцину.
Меланоцитстимулирующие гормоны — гормоны средней, или промежуточной, доли гипофиза позвоночных животных и человека. Стимулирует синтез и секрецию меланинов (меланогенез) клетками-меланоцитами кожи и волос, а также пигментного слоя сетчатки глаза.
7. В надпочечниках выделяют корковое и мозговое вещество. Корковое вещество включает клубочковую, пучковую и сетчатую зоны. В клубочковой зоне происходит синтез минералокортикоидов, основным представителем которых является альдостерон. В пучковой зоне синтезируются глюкокортикоиды. В сетчатой зоне вырабатывается небольшое количество половых гормонов.
Альдостерон усиливает в дистальных канальцах почек реабсорбцию ионов Na+, одновременно увеличивая при этом выведение-с мочой ионов К+. Кроме того, под влиянием альдостерона резко возрастает почечная реабсорбция воды. Альдостерон является провоспалительным гормоном. Под влиянием альдостерона увеличивается также секреция ионов Н+в канальцевом аппарате почек, что приводит к снижению их концентрации во внеклеточной жидкости и изменению кислотно-основного состояния (алкалоз).
Основным фактором, регулирующим секрецию альдостерона, является функционирование ренин-ангиотензин-альдостероновой системы. При снижении уровня АД наблюдается возбуждение симпатической части автономной нервной системы, что приводит к сужению почечных сосудов. Уменьшение почечного кровотока способствует усиленной выработке ренина в юкстагломерулярных нефронах почек. Ренин является ферментом, который действует на плазменный α 2-глобулин ангиотензиноген, превращая его в ангиотензин I. Образовавшийся ангиотензин I затем превращается в ангиотензин II, который увеличивает секрецию альдостерона. Выработка альдостерона может усиливаться также по механизму обратной связи при изменении электролитного состава плазмы крови, в частности при гипонатриемии или гиперкалиемии. В незначительной степени секреция этого гормона стимулируется кортикотропином.
Глюкокортикоиды вызывают следующие эффекты:
1. Влияют на все виды обмена веществ:
а) на белковый обмен. Под влиянием глюкокортикоидов стимулируются процессы распада белка.
б) на жировой обмен. Глюкокортикоиды усиливают мобилизацию жира из жировых депо и увеличивают концентрацию жирных кислот в плазме крови. Вместе с тем увеличивается отложение жира в области лица, груди и на боковых поверхностях туловища.
в) на углеводный обмен. Введение глюкокортикоидов приводит к увеличению содержания глюкозы в плазме крови (гипергликемия). В основе этого эффекта лежит стимулирующее действие на процессы глюконеогенеза. Избыток аминокислот, образовавшихся в результате катаболизма белка, используется для синтеза глюкозы в печени. Гипергликемический эффект является одним из компонентов защитного действия глюкокортикоидов при стрессе.
2. Противовоспалительное действие. Глюкокортикоиды угнетают все стадии воспалительной реакции (альтерацию, экссудацию и пролиферацию), стабилизируют мембраны лизосом, что предотвращает выброс протеолитических ферментов, способствующих развитию воспалительной реакции.
3. Противоаллергическое действие. Гиперпродукция глюкокортикоидов приводит к снижению числа эозинофилов в крови, увеличенное количество которых обычно является «маркером» аллергии.
4. Подавление иммунитета. Глюкокортикоиды угнетают как клеточный, так и гуморальный иммунитет, что связано со снижением образования антител и процессов фагоцитоза. Длительный прием глюкокортикоидов приводит к инволюции тимуса и лимфоидной ткани, являющихся иммунокомпетентными органами, вследствие чего уменьшается количество лимфоцитов в крови. Подавление иммунитета может являться серьезным побочным эффектом при длительном приеме глюкокортикоидов, поскольку при этом возрастает вероятность присоединения вторичной инфекции. С другой стороны, этот эффект может являться терапевтическим при использовании глюкокортикоидов для подавления роста опухолей, происходящих из лимфоидной ткани, или для торможения реакций отторжения при трансплантации органов и тканей.
5. Участие в формировании необходимого уровня АД. Глюкокортикоиды повышают чувствительность сосудистой стенки к действию катехоламинов, что приводит к гипертензии.
В организме существует определенный суточный ритм выработки глюкокортикоидов. Основная масса этих гормонов вырабатывается в утренние часы (6—8 ч утра). Последнее учитывают при распределении суточной дозы гормонов в процессе длительного лечения глюкокортикоидами.
Продукция глюкокортикоидов регулируется кортикотропином. Его выделение усиливается при действии на организм стрессорных стимулов различной природы, что является пусковым моментом для развития адаптационного синдрома.
Половые гормоны. При избыточном образовании половых гормонов в сетчатой зоне развивается адреногенитальный синдром двух типов — гетеросексуальный и изосексуальный. Гетеросексуальный синдром развивается при выработке гормонов противоположного пола и сопровождается появлением вторичных половых признаков, присущих другому полу. Изосексуальный синдром наступает при избыточной выработке гормонов одноименного пола и проявляется ускорением процессов полового развития.
8. Катехоламины. В мозговом веществе надпочечников содержатся хромаффинные клетки, в которых синтезируются адреналин и норадреналин. Примерно 80% гормональной секреции приходится на адреналин и 20% — на норадреналин. Продукция этих гормонов резко усиливается при возбуждении симпатической части автономной нервной системы. В свою очередь выделение этих гормонов в кровь приводит к развитию эффектов, аналогичных действию стимуляции симпатических нервов. Разница состоит лишь в том, что гормональный эффект является более длительным. К наиболее важным эффектам катехоламинов относятся стимуляция деятельности сердца, вазоконстрикция, торможение перистальтики и секреции кишечника, расширение зрачка, уменьшение потоотделения, усиление процессов катаболизма и образования энергии. Адреналин имеет большее сродство к β -адренорецепторам, локализующимся в миокарде, вследствие чего вызывает положительные инотропный и хронотропный эффекты в сердце. С другой стороны, норадреналин имеет более высокое сродство к сосудистым α -адренорецепторам. Поэтому, вызываемые катехоламинами вазоконстрикция и увеличение периферического сосудистого сопротивления в большей степени обусловлены действием норадреналина.
Вопросы для самоподготовки
- Общие представления об эндокринной системе. Определение гормонов. Классификация, типы действия и функции гормонов.
- Механизмы действия гормонов.
- Регуляция концентрации гормонов в крови и тканях.
- Гипоталамус. Гипоталамо-гипофизарная система
- Гипофиз. Гормоны гипофиза. Физиологическое действие гормонов нейрогипофиза. Меланоцитостимулирующий гормон.
- Гормоны аденогипофиза и их функции.
- Гормоны коры надпочечников и их физиологическое действие.
- Симпатоадреналовая система (гормоны, место выработки, регуляция секреции, эффекты действий).
|
|