Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Хромосомная теория наследственности,
|
|
теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности, т.е. преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом. Х. т. н. возникла в начале 20 в. на основе клеточной теории и использования для изучения наследственных свойств организмов гибридологического анализа.
В 1902 У. Сеттон в США, обративший внимание на параллелизм в поведении хромосом и менделевских т. н. " наследственных факторов", и Т. Бовери в Германии выдвинули хромосомную гипотезу наследственности, согласно которой менделевские наследственные факторы (название впоследствии генами) локализованы в хромосомах. Первые подтверждения этой гипотезы были получены при изучении генетического механизма определения пола у животных, когда было выяснено, что в основе этого механизма лежит распределение половых хромосом среди потомков. Дальнейшее обоснование Х. т. н. принадлежит американскому генетику Т. Х. Моргану, который заметил, что передача некоторых генов (например, гена, обусловливающего белоглазие у самок дрозофилы при скрещивании с красноглазыми самцами) связана с передачей половой Х-хромосомы, т. е. что наследуются признаки, сцепленные с полом (у человека известно несколько десятков таких признаков, в том числе некоторые наследственные дефекты — дальтонизм, гемофилия и др.).
Доказательство Х. т. н. было получено в 1913 американским генетиком К. Бриджесом, открывшим нерасхождение хромосом в процессе мейоза у самок дрозофилы и отметившим, что нарушение в распределении половых хромосом сопровождается изменениями в наследовании признаков, сцепленных с полом.
С развитием Х. т. н. было установлено, что гены, расположенные в одной хромосоме, составляют одну группу сцепления и должны наследоваться совместно; число групп сцепления равно числу пар хромосом, постоянному для каждого вида организмов, признаки, зависящие от сцепленных генов, также наследуются совместно. Вследствие этого закон независимого комбинирования признаков должен иметь ограниченное применение; независимо должны наследоваться признаки, гены которых расположены в разных (негомологичных) хромосомах. Явление неполного сцепления генов (когда наряду с родительскими сочетаниями признаков в потомстве от скрещиваний обнаруживаются и новые, рекомбинантные, их сочетания) было подробно исследовано Морганом и его сотрудниками (А. Г. Стёртевантом и др.) и послужило обоснованием линейного расположения генов в хромосомах. Морган предположил, что сцепленные гены гомологичных хромосом, находящиеся у родителей в сочетаниях  и
и  , в мейозе у гетерозиготной формы
, в мейозе у гетерозиготной формы 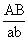 могут меняться местами, в результате чего наряду с гаметами АВ и ab образуются гаметы Ab и аВ. Подобные перекомбинации происходят благодаря разрывам гомологичных хромосом на участке между генами
могут меняться местами, в результате чего наряду с гаметами АВ и ab образуются гаметы Ab и аВ. Подобные перекомбинации происходят благодаря разрывам гомологичных хромосом на участке между генами  и последующему соединению разорванных концов в новом сочетании: Реальность этого процесса, названного перекрестом хромосом, или кроссинговером, была доказана в 1933 нем, учёным К. Штерномв опытах с дрозофилой и американскими учёными Х. Крейтономи Б. Мак-Клинток — с кукурузой. Чем дальше друг от друга расположены сцепленные гены, тем больше вероятность кроссинговера между ними. Зависимость частоты кроссинговера от расстояний между сцепленными генами была использована для построения генетических карт хромосом. В 30-х гг. 20 в. Ф. Добржанский показал, что порядок размещения генов на генетических и цитологических картах хромосом совпадает.
и последующему соединению разорванных концов в новом сочетании: Реальность этого процесса, названного перекрестом хромосом, или кроссинговером, была доказана в 1933 нем, учёным К. Штерномв опытах с дрозофилой и американскими учёными Х. Крейтономи Б. Мак-Клинток — с кукурузой. Чем дальше друг от друга расположены сцепленные гены, тем больше вероятность кроссинговера между ними. Зависимость частоты кроссинговера от расстояний между сцепленными генами была использована для построения генетических карт хромосом. В 30-х гг. 20 в. Ф. Добржанский показал, что порядок размещения генов на генетических и цитологических картах хромосом совпадает.
Согласно представлениям школы Моргана, гены являются дискретными и далее неделимыми носителями наследственной информации. Однако открытие в 1925 советскими учёными Г. А. Надсоном и Г. С. Филипповым, а в 1927 американским учёным Г. Мёллером влияния рентгеновских лучей на возникновение наследственных изменений (мутаций) у дрозофилы, а также применение рентгеновских лучей для ускорения мутационного процесса у дрозофилы позволили советским учёным А. С. Серебровскому, Н. П. Дубинину и др. сформулировать в 1928—30 представления о делимости гена на более мелкие единицы, расположенные в линейной последовательности и способные к мутационным изменениям. В 1957 эти представления были доказаны работой американского учёного С. Бензера с бактериофагом Т4. Использование рентгеновских лучей для стимулирования хромосомных перестроек позволило Н. П. Дубинину и Б. Н. Сидорову обнаружить в 1934 эффект положения гена (открытый в 1925 Стёртевантом), т. е. зависимость проявления гена от места расположения его на хромосоме. Возникло представление о единстве дискретности и непрерывности в строении хромосомы.
Х. т. н. развивается в направлении углубления знаний об универсальных носителях наследственной информации — молекулах дезоксирибонуклеиновой кислоты (ДНК). Установлено, что непрерывная последовательность пуриновых и пиримидиновых оснований вдоль цепи ДНК образует гены, межгенные интервалы, знаки начала и конца считывания информации в пределах гена; определяет наследственный характер синтеза специфических белков клетки и, следовательно, наследственный характер обмена веществ. ДНК составляет материальную основу группы сцепления у бактерий и многих вирусов (у некоторых вирусов носителем наследственной информации является рибонуклеиновая кислота); молекулы ДНК, входящие в состав митохондрий, пластид и др. органоидов клетки, служат материальными носителями цитоплазматической наследственности.
Х. т. н., объясняя закономерности наследования признаков у животных и растительных организмов, играет важную роль в с.-х. науке и практике. Она вооружает селекционеров методами выведения пород животных и сортов растений с заданными свойствами. Некоторые положения Х. т. н. позволяют более рационально вести с.-х. производство. Так, явление сцепленного с полом наследования ряда признаков у с.-х. животных позволило до изобретения методов искусственного регулирования пола у тутового шелкопряда выбраковывать коконы менее продуктивного пола, до разработки способа разделения цыплят по полу исследованием клоаки — отбраковывать петушков и т.п. Важнейшее значение для повышения урожайности многих с.-х. культур имеет использование полиплоидии. На знании закономерностей хромосомных перестроек основывается изучение наследственных заболеваний человека.
Генетические карты хромосом, схемы относительного расположения сцепленных между собой наследственных факторов — генов. Имея Г. к. х., можно по наследованию " сигнального" гена, тесно сцепленного с изучаемым, контролировать передачу потомству генов, обусловливающих развитие трудно анализируемых признаков; например, ген, определяющий эндосперм у кукурузы и находящийся в 9-й хромосоме, сцеплен с геном, определяющим пониженную жизнеспособность растения. Многочисленные факты отсутствия (вопреки Менделя законам) независимого распределения признаков у гибридов второго поколения были объяснены хромосомной теорией наследственности. Гены, расположенные в одной хромосоме, в большинстве случаев наследуются совместно и образуют одну группу сцепления, количество которых, т. о., соответствует у каждого организма гаплоидному числу хромосом. Американский генетик Т. Х. Морган показал, однако, что сцепление генов, расположенных в одной хромосоме, у диплоидных организмов не абсолютное; в некоторых случаях перед образованием половых клеток между однотипными, или гомологичными, хромосомами происходит обмен соответственными участками; этот процесс носит название перекреста, или кроссинговера. Обмен участками хромосом (с находящимися в них генами) происходит с различной вероятностью, зависящей от расстояния между ними (чем дальше друг от друга гены, тем выше вероятность кроссинговера и, следовательно, рекомбинации). Генетический анализ позволяет обнаружить перекрест только при различии гомологичных хромосом по составу генов, что при кроссинговере приводит к появлению новых генных комбинаций. Обычно расстояние между генами на Г. к. х. выражают как % кроссинговера (отношение числа мутантных особей, отличающихся от родителей иным сочетанием генов, к общему количеству изученных особей); единица этого расстояния — морганида — соответствует частоте кроссинговера в 1%.
Г. к. х. составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, по мере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые названия мутантных генов, их расстояния в морганидах от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры. Составить Г. к. х. можно только для объектов, у которых изучено большое число мутантных генов. Например, у дрозофилы идентифицировано свыше 500 генов, локализованных в её 4 группах сцепления, у кукурузы — около 400 генов, распределенных в 10 группах сцепления. У менее изученных объектов число обнаруженных групп сцепления меньше гаплоидного числа хромосом. Так, у домовой мыши выявлено около 200 генов, образующих 15 групп сцепления (на самом деле их 20); у кур изучено пока всего 8 из 39. У человека из ожидаемых 23 групп сцепления (23 пары хромосом) идентифицировано только 10, причём в каждой группе известно небольшое число генов; наиболее подробные карты составлены для половых хромосом. У бактерий, которые являются гаплоидными организмами, имеется одна, чаще всего непрерывная, кольцевая хромосома и все гены образуют одну группу сцепления. При переносе генетического материала из клетки-донора в клетку-реципиент, например при конъюгации, кольцевая хромосома разрывается и образующаяся линейная структура переносится из одной бактериальной клетки в другую (у кишечной палочки в течение 110—120 мин). Искусственно прерывая процесс конъюгации, можно по возникшим типам рекомбинантов установить, какие гены успели перейти в клетку-реципиент. В этом состоит один из методов построения Г. к. х. бактерий, детально разработанных у ряда видов. Ещё более детализированы Г. к. х. некоторых бактериофагов.
21. Кариотип и идиограмма хромосом человека. Денверская и Парижская классификации хромосом. Характеристика кариотипа человека в норме и при патологии.
Кариоти́ п — совокупность признаков (число, размеры, форма и т.д.) полного набора хромосом, присущий клеткам данного биологического вида (видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Кариотипом иногда также называют и визуальное представление полного хромосомного набора (кариограммы).
|
|