Главная страница Случайная страница
Разделы сайта
АвтомобилиАстрономияБиологияГеографияДом и садДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеталлургияМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРелигияРиторикаСоциологияСпортСтроительствоТехнологияТуризмФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Тема 3. Популяція та її основні характеристики
|
|
Особини одного виду зустрічаються на територіях, де вони знаходять придатні для життя умови. Частину земної поверхні (чи акваторії), в межах якої зустрічається даний вид, називається ареалом (від лат. «area» – площа, простір).
Розміри ареалів різних видів можуть сильно відрізнятися. У наземних малорухомих видів, поширення яких обмежено якимись непереборними перешкодами, ареал може займати територію всього в кілька квадратних кілометрів і навіть менше. До них відносяться острівні або печерні види, мешканці гірських долин або верхніх зон гірських хребтів. Види, що мають вузький ареал поширення, називаються ендемічними.
Інші види мають великі ареали, нерідко розташовані на декількох материках. Наприклад, на всіх континентах зустрічаються жуки-гнойовики, соколи-сапсани. Величезний ареал мають рухливі морські тварини – кашалот, сірий дельфін, синій кит, косатка. Таке ж широке поширення властиво багатьом рослинам і тваринам, які супроводжують людину, - синантропним видами (воші, блохи, таргани, щури). Види, ареали яких розташовані в межах усіх континентів, називаються світовими, або космополітами.
Головними причинами, які впливають на формування та особливості структури ареалу, є екологічна пластичність виду, його здатність до розселення та історичний вік. У формуванні ареалів синантропних видів роль людини є визначальною.
Популяція – структурна одиниця виду. Ареалів, суцільно заселених тим чи іншим видом, в природі не існує. В межах ареалу особини даного виду освоюють лише відповідне для їх життя місцеперебування. Ступінь заповнення займаного простору у різних видів різна. Але завжди в ньому виділяються «порожнечі» і скупчення. Іншими словами, ареал складається з більш або менш численних ділянок, на яких і зустрічається певний вид. Наприклад, колонії крота європейського, добре помітні по горбкам землі, розташовуються на лісових галявинах і луках, ялина звичайна росте переважно на низинах зі значно зволоженим ґрунтом.
Скупчення особин одного виду за чисельністю можуть бути великими або маленькими, існувати довгостроково (століття і більше) або протягом життя двох-трьох поколінь, після чого вони, як правило, гинуть від будь-яких випадковостей, наприклад, захворювань, різкого погіршення погодних умов та ін.
Для долі виду набагато більш важливу роль відіграють ті групи особин, які стабільно зберігаються протягом життя багатьох поколінь. Чисельність особин в таких групах може значно збільшуватися при сприятливих умовах і знижуватися при несприятливих, однак вони мають шанси на тривале існування на даній території. Такі угруповання (сукупності) особин одного виду, які тривалий населяють певну частину ареалу, які вільно схрещуються один з одним і дають плодюче потомство, відносно відокремлені від інших сукупностей того ж виду, називається популяцією (від лат. populus – народ, населення).
Завдяки просторової розрізненості популяцій вид пристосований до існування в різноманітних умовах середовища. Таким чином, популяція є внутрішньовидовим угрупованням і, отже, конкретною формою існування виду, а сам вид – складною біологічною системою.
Кожна популяція будь-якого виду як біологічна система володіє певною структурою. Під структурою популяції розуміється певне кількісне співвідношення особин, які відрізняються за морфологічними і фізіологічними ознаками, віком, статтю, характером розподілу у просторі та іншим властивостям.
Основними параметрами популяції є, насамперед, її чисельність і щільність.
Чисельність – загальна кількість особин в популяції. Вона не буває постійною, так як мінливі умови середовища існування популяції. Чисельність популяції залежить від співвідношення інтенсивності розмноження (плодючості) і смертності. В процесі розмноження відбувається зростання популяції, смертність ж призводить до скорочення її чисельності. Для кожної популяції є верхній і нижній межі чисельності, які можна виміряти, вивчаючи її сезонні та міжрічні зміни.
Щільність популяції – це кількість особин або їх біомаса на одиницю площі або об'єму (наприклад, 150 рослин сосни на 1 га; 0, 5 циклопа на 1 м3 води). Щільність популяції також мінлива і залежить від чисельності. При зростанні чисельності щільність не збільшується лише в тому разі, якщо можливо розселення популяції, розширення її ареалу.
Підвищення щільності понад оптимальної несприятливо позначається на стані популяції, оскільки при цьому вичерпується кормова база, скорочується життєвий простір і т. д.
Падіння щільності нижче оптимальної призводить до ослаблення захисних реакцій популяції, знижує її плодючість, що в кінцевому підсумку може призвести до вимирання популяції.
Народжуваність характеризує частоту появи нових особин в популяції за рахунок розмноження. Розрізняють абсолютну і питому народжуваність.
Абсолютна народжуваність – це кількість особин, народжених у популяції за одиницю часу, наприклад, за рік, місяць, добу. Однак величина абсолютної народжуваності знаходиться в прямій залежності від чисельності популяції. Тому визначають питому народжуваність – кількість особин, народжених у популяції за одиницю часу в розрахунку на одну особину (або на тисячу особин). Показник питомої народжуваності зручний тим, що дозволяє порівнювати швидкість народжуваності в різних популяціях незалежно від відмінностей у їх чисельності.
Смертність (абсолютна і відносна) – величина, протилежна народжуваності – число померлих або загиблих особин (m) по відношенню до їх загального числа (N) в популяції (частіше на 100 або 1000 особин) на певній території; інтенсивність процесу загибелі особин в популяції.
Величини народжуваності і смертності мінливі в часі і залежать від вікової і статевої структури популяції, забезпеченості їжею і ряду інших факторів. Співвідношення між величинами народжуваності і смертності визначає динаміку чисельності популяції. Так, якщо величина народжуваності вище показника смертності, то чисельність популяції зростатиме, і навпаки, знизиться, якщо смертність перевищуватиме народжуваність. У разі рівності величин народжуваності і смертності чисельність популяції буде підтримуватися на постійному рівні.
У природних умовах зміна чисельності популяції носить коливальний характер. Амплітуда коливань залежить від особливостей виду і умов існування. У багатьох великих хребетних (при відносно постійних умовах навколишнього середовища) чисельність коливається лише в кілька разів, тоді як, наприклад, у комах, коливання чисельності популяції ширше – в 40-50 разів, а в окремі роки, за сприятливих умов, виникають різкі спалахи чисельності, коли вона зростає в десятки і навіть у мільйони разів. Наприклад, спалахи чисельності сарани, що відбуваються з інтервалом приблизно в сорок років.
Крім нерегулярних спалахів чисельності, можуть мати місце і регулярні спалахи, які будуть обумовлені різними факторами середовища (температура, харчові ресурси тощо).
Фактори, що впливають на чисельність популяції, ділять на не залежні (абіотичні фактори) і залежні (біотичні фактори – конкуренція, хижацтво, забезпеченість їжею, поширення інфекцій і т. д.) від її щільності. Множинність механізмів регуляції чисельності популяції в природних умовах сприяє тому, що в природних популяціях нечасто спостерігається катастрофічне зростання чисельності, підрив ресурсів і загибель популяції.
Всі пристосувальні реакції організмів у природі спрямовані на те, щоб підвищити ймовірність виживання і залишення потомства.
Загальну чисельність популяції оцінюють різними методами. Для підрахунку популяцій великорозмірних організмів, таких як слони, буйволи, олені, застосовують метод тотального підрахунку, використовуючи фотозйомку території з літака або гелікоптера з подальшим прораховуванням на фотографії всіх зображених на ній особин даного виду.
Чисельність популяцій, які складаються з дрібних тварин, найчастіше визначають методом вибірки. Для цього підраховують щільність організмів в різних ділянках місцеперебування популяції і, помноживши середню величину щільності на площу або об'єм, займаний популяцією, отримують її загальну чисельність.
Просторовий розподіл являє собою особливості розміщення особин популяції на займаній території. Воно визначається ступенем однорідності середовища проживання, наявністю придатних для життя ділянок, а також біологічними особливостями виду, поведінкою його особин. Знання типу розподілу організмів дозволяє правильно оцінити щільність методом вибірки.
Природним популяціям властиві три типи розподілу особин: випадковий (дифузний), рівномірний (регулярний) і груповий (сукупний).
Випадковий (дифузний) розподіл особин спостерігається в однорідному середовищі, при невисокій чисельності популяції і відсутності у особин прагнення утворювати групи (наприклад, у планарій, гідр). У природі цей тип розподілу зустрічається нечасто.
Рівномірний (регулярний) розподіл характерний для видів, що відрізняються жорсткою конкуренцією між особинами за однакові ресурси і сильним територіальним інстинктом (хижі риби, ссавці, птахи, павуки).
Груповий (сукупний) розподіл зустрічається в природі найбільш часто. Він висловлюється в утворенні угруповань особин, між якими залишаються значні незаселені території. Причинами аґрегованості особин можуть бути неоднорідність середовища і обмеженість придатних для життя місцезростань, особливості розмноження, прагнення до життя в групі. Життя у групі (сім'ї, стаді, зграї, колонії) полегшує захист від хижаків, пошук та добування корму. У рослин груповий розподіл визначається головним чином способом розмноження і розселення насіння і плодів. Наприклад, важкі плоди, падаючи поряд з материнською рослиною і в подальшому проростаючи, утворюють групу поруч зростаючих дочірніх рослин.
Вікова структура відображає співвідношення різних вікових груп у популяції, а також сезонну і міжрічну динаміку цього співвідношення. В популяції зазвичай виділяють три екологічних віка: передрепродуктивний (до розмноження), репродуктивний (у період розмноження) і післяпродуктивний (після розмноження).
При сприятливих умовах у популяції присутні всі вікові групи і підтримується більш або менш стабільний рівень її чисельності. Незважаючи на присутність в популяції всіх вікових груп, співвідношення між ними змінюється і залежить від числа нащадків в посліді, кількості приплодів за сезон, часу настання статевозрілості, тривалості репродуктивного періоду особин, тривалості їх життя та інших факторів. Так, наприклад, у популяціях дрібних гризунів велика частка особин молодших вікових груп, в той час як для великих видів (слони, кити та ін.) вона мала.
У зменшенні популяціях переважають старі особини, вже не здатні інтенсивно розмножуватися. Така вікова структура свідчить про несприятливі умови існування.
Вивчення розподілу організмів за віком має велике значення в прогнозуванні чисельності популяцій протягом життя ряду найближчих поколінь. Такі дослідження дозволяють планувати, наприклад, промисел риб або хутрових звірів на кілька років вперед.
Статеву структуру формує співвідношення статей у популяціях з роздільностатевими особинами. До них відносяться більшість тварин та дводомні рослини. Зміна статевої структури популяції відбивається на її ролі в екосистемі, так як самці і самки багатьох видів мають відмінності в характері харчування, ритмі життя, поведінці. Так, самки деяких видів комарів, кліщів і комах є кровопивцями, в той час як самці живляться соком рослин або нектаром. Підвищення частки статевозрілих самок призводить до більш інтенсивному росту популяції за рахунок народження більшої кількості молоді.
Етологічна структура популяцій – це система відносин між членами однієї популяції. Форми спільного існування особин у популяціях різноманітні.
Одиночний спосіб життя, при якому особини популяції незалежні і відокремлені один від одного, характерний для багатьох видів, але лише на певних стадіях життєвого циклу. Повністю одиночне існування організмів у природі не зустрічається, так як при цьому було б неможливим здійснення їх основної життєвої функції – розмноження. Однак для деяких видів характерні дуже слабкі контакти між спільно живуть особинами. Такими є, зокрема, окремі водні мешканці з зовнішнім способом запліднення, при якому немає необхідності в безпосередній зустрічі партнерів, наприклад поодинокі актинії. У видів з внутрішнім заплідненням зустрічі самців і самок також можуть бути дуже короткочасними, лише для здійснення копуляції, в інший час тварини живуть незалежно один від одного. Такий спосіб життя багатьох комах, наприклад сонечка, хижих жуків-турунів та ін.
У видів з одиночним способом життя часто утворюються тимчасові скупчення особин у місцях зимівлі, у період, передуючий розмноженню, і т. п. Так, метелики-кропив'янки пізньої осені цілими гронами збираються в горищних приміщеннях або інших укриттях, жужелиці і сонечка – в сухій підстилці біля пнів і коренів дерев, соми та щуки – в зимувальних ямах на дні водойми. У бентосних тварин такі скупчення можуть бути постійними, наприклад устричні або мідієві банки, агрегації поліхет або поселення морських жолудів. У цих поселеннях збільшуються шанси на можливість запліднення статевими продуктами, викидаються у воду. Однак подібні агрегації не супроводжуються встановленням більш або менш закономірного поведінки тварин по відношенню одне до одного, і кожне з них відносно незалежно від інших.
Подальше ускладнення відносин всередині популяції здійснюється за двома напрямами: посилення зв'язку між статевими партнерами й виникнення контактів між батьківським і дочірнім поколіннями. На цій основі в популяціях формуються родини, які дуже різноманітні за складом і тривалості існування.
Сім'я – група особин, в якій посилюється зв'язок між нащадками і батьками. Вона може бути змішаного типу (птахи, деякі ссавці); батьківського типу (виховання здійснює самець, наприклад африканський страус, риба колюшка) материнського типу (саламандра, черв'яга (безноге земноводне).
Батьківські пари можуть створюватися як на короткий, так і на тривалий термін, а у деяких видів – на все життя дорослих особин. Наприклад, серед птахів полігамні тетерева, глухарі спаровуються з багатьма самками, не утворюючи стійких сімейних пар. У деяких качок – крижні, шилохвости та ін. – пари підбираються ще на місцях зимівлі або під час перельоту, але, після того, як самка сідає на гніздо, самець її покидає. У більшості горобцевих самець і самка тримаються разом протягом всього періоду гніздування. Сімейні пари лебедів, журавлів, голубів зберігаються на довгі роки.
Розрізняють сім'ї батьківського, материнського і змішаного типу, в залежності від того, хто з батьків бере на себе догляд за потомством. У сім'ях з стійким утворенням пар в охороні та вигодовуванні молодняку зазвичай приймають участь і самець і самка.
При сімейному способі життя територіальна поведінка тварин виражена найбільш яскраво: різні сигнали, маркування, ритуальні форми загрози і пряма агресія забезпечують володіння ділянкою, достатнім для вигодовування потомства.
Колонія – це групове об'єднання осілих тварин. Вони можуть існувати довго або виникати на період розмноження (птахи, морські котики, тюлені, терміти, бджоли, мурашки, бабаки, лемінги).
За складністю взаємозв'язків між особинами колонії тварин надзвичайно різноманітні – від простих територіальних скупчень одиночних форм до об'єднань, де окремі члени виконують, як органи в цілісному організмі, різні функції видовий життя.
Поселення морських жолудів, наприклад, можна розглядати і як агрегації поодиноких особин, і як елементарну форму колонії, тому що одновидові скупчення виникають у них не випадково, а на основі хемотаксису, властивого личинкам. Взаємовідносини між особинами в таких примітивних об'єднаннях зводяться до стимуляції синхронного статевого дозрівання хімічними виділеннями членів колонії, а конкуренція за простір для лову певною мірою припиняється орієнтовною реакцією личинок перед прикріпленням.
Колонії ссавців частіше виникають не як територіальні об'єднання різних сімей, а на основі розростання сімейних груп, із збереженням зв'язків між сім'ями, які відмежовуються. Тварин з інших колоній при повній зайнятості території виганяють, але в період низької чисельності популяції вони можуть вливатися в склад поселення, яке, таким чином, набуває змішаний характер.
Найбільш складні колонії у громадських комах – термітів, мурашок, бджіл. Вони виникають на основі сильно розростається сім'ї. В таких колоніях-сім'ях комахи виконують спільно більшість основних функцій: розмноження, захисту, забезпечення кормом себе і потомства, будівництва і т. п. При цьому існує обов'язковий поділ праці та спеціалізація окремих особин або вікових груп на виконанні певних операцій. Члени колонії діють на основі постійного обміну інформацією один з одним.
Зграя – це тимчасове об'єднання тварин одного виду (вовки, птахи, риби). Зграї полегшують виконання будь-яких функцій в житті виду: захисту від ворогів, добування їжі, міграції. Найбільш широко зграйність поширена серед птахів і риб, ссавців характерна для багатьох собачих. У зграях сильно розвинені наслідувальні реакції і орієнтація на сусідів.
За способами координації дій зграї діляться на дві категорії: еквіпотенціальні, без вираженого домінування окремих членів і зграї з лідерами, в яких тварини орієнтуються на поведінку однієї або кількох, зазвичай найбільш досвідчених, особин. Об'єднання першого типу характерні в основному для риб, але відомі також у дрібних птахів, перелітної сарани і деяких інших форм. Другий тип зграй зустрічається зазвичай у великих птахів і ссавців.
У зграї ссавців велику роль відіграють ватажки, специфічні відносини складаються між окремими особинами, що зближує ці групові утворення зі стадами.
Стада – тривалі і постійне об'єднання тварин в порівнянні зі зграями. У стадних групах, як правило, здійснюються всі основні функції життя виду: добування корму, захист від хижаків, міграції, розмноження, виховання молодняку і т. п. Основу групової поведінки тварин у стадах складають взаємовідносини домінування-підпорядкування, засновані на індивідуальних відмінностях між особинами.
Один з варіантів організації стад – це групи з тимчасовими або постійними лідерами – особинами, на яких концентрується увага інших і які своєю поведінкою визначають напрямок переміщення, місця годівлі, реакцію на хижаків та інші властивості стада. Стадо діє як єдине ціле, наслідуючи лідеру. Діяльність лідера не спрямована безпосередньо на підпорядкування інших особин. Лідером стає більш досвідчений член стада.
Система домінування особин у популяціях може бути різною, а саме:
1) ієрархія за схемою «трикутника»: А нападає на В, В – С, а С підпорядковує собі А;
2) лінійна ієрархія в ряду рангів А – В – С (особини наступного рангу підпорядковані попередньому). Подібні відносини виникають у зграях їздових собак;
3) паралельна ієрархія (окремо серед самців та серед самок); наприклад, у популяціях мавп;
4) деспотія (домінування однієї особи над усіма іншими – мавпи).
Біологічне значення відносин між особинами одного виду: захист тварин від ворогів; виховання молоді; отримання їжі і т. д.
Екологічна стратегія – загальна характеристика росту і розмноження даного виду. Вона включає темп росту особин, час досягнення статевої зрілості, плодючість і інші характеристики. Екологічна стратегія залежить від багатьох факторів, але особливо важливі ті, які впливають на стратегію росту і розмноження, тобто фактори, що впливають на смертність.
Екологічні стратегії вельми різноманітні, але виділяють в основному два крайніх типу стратегії, які отримали назву r - і К-стратегії (символи взяті з рівняння логістичного зростання чисельності популяції). r-стратегія визначається відбором на підвищення швидкості росту популяції в періоди її низької щільності, а К-стратегія виникає під дією відбору на підвищення виживаності при високій щільності популяції, близькою до ємності середовища.
К-стратеги повільно розвиваються, мають більш великі розміри і велику тривалість життя, утворюють невелике число великих, добре захищених нащадків; r-стратегія частіше зустрічається у організмів, що живуть у середовищі з різкими змінами умов середовища – в ефемерних (існуючих дуже недовго) біотопах, в пересихаючих калюжах. Чисельність в таких популяціях різко коливається, але спостерігається висока швидкість розмноження, так як сприятливий період часу дуже недовгий і особинам вигідно швидко досягати статевозрілості. K-стратегія характерна більше для популяцій, що мешкають в стабільних умовах середовища або в умовах, які закономірно змінюються (наприклад, сезонні зміни). Тому у k-стратегів потомство менш численне, але зате конкурентноздатне. Між r - і k-стратегіями багато переходів, але виділення цих двох основних типів стратегій допомагає пояснити зв'язок між різноякісними характеристиками популяції та умовами середовища.
Екологічна ніша – місце виду в біогеоценозі, визначається його біотичних потенціалом і сукупністю факторів зовнішнього середовища, до яких він пристосований. Це не тільки фізичний простір, зайнятий організмом, але і його функціональна роль в співтоваристві (положення в харчовому ланцюзі), і його місце щодо зовнішніх факторів.
У структурі екологічної ніші виділяють 3 складові:
Просторова ніша (місце проживання) – " адреса" організму;
Трофічна ніша – характерні особливості харчування і роль виду в співтоваристві – " професія";
Багатовимірна (гіперпросторова) екологічна ніша – діапазон всіх умов, за яких живе і відтворює себе особина або популяція.
Розрізняють фундаментальну (потенційну) нішу, яку організм або вид міг би займати в відсутність конкуренції, хижаків, в якій абіотичні умови оптимальні; і реалізовану нішу – фактичний діапазон умов існування організму, який менше або дорівнює фундаментальної ніші.
Правило обов'язковості заповнення екологічної ніші – порожня екологічна ніша завжди і обов'язково буває природно заповнена. В насичених біогеоценозах життєві ресурси використовуються найбільш повно – у них всі екологічні ніші зайняті. У ненасичених біогеоценозах життєві ресурси частково утилізуються, для них характерно наявність вільних екологічних ніш.
Екологічне дублювання – захоплення звільненої екологічної ніші іншим видом, здатним виконувати в співтоваристві ті ж функції, що і зниклий вид. З цього випливає, що, знаючи розподіл видів по екологічній ніші в співтоваристві і параметри кожної екологічної ніші, можна заздалегідь описати вид, який зможе зайняти ту чи іншу нішу у випадку її звільнення.
Екологічна диверсифікація – явище поділу екологічної ніші в результаті міжвидової конкуренції. Здійснюється за трьома параметрами:
- за просторовим розміщенням;
- за харчовим раціоном;
- за розподілом активності в часі.
Внаслідок диверсифікації відбувається зміщення ознак – особини двох близьких видів більш подібні між собою в тих частинах ареалів, де зустрічаються окремо, ніж на ділянках спільного проживання.
Характеристики екологічної ніші: ш ирина і перекривання даної ніші сусідніми.
Ширина екологічної ніші – відносний параметр, який оцінюють шляхом порівняння з шириною екологічної ніші інших видів. Еврибіонти зазвичай мають більш широкі екологічні ніші, ніж стенобіонти. Однак, одна і та ж екологічна ніша може мати різну ширину за різними напрямками: наприклад, за просторовим розподілом, харчових зв'язкам і т. д.
Перекривання екологічної ніші виникає, якщо різні види при спільному проживанні, використовують одні і ті ж ресурси. Перекривання може бути повним або частковим, по одному або декільком параметрам екологічної ніші.
Якщо екологічна ніша одного виду включає в себе екологічну нішу іншого (рис.1), то виникає інтенсивна конкуренція, домінуючий конкурент витіснить свого суперника на периферію зони пристосованості.

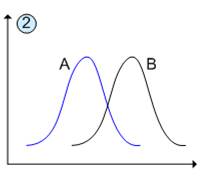
Якщо екологічні ніші частково перекриваються (рис.2), то їх спільне співіснування буде можливо завдяки наявності у кожного виду специфічних пристосувань.
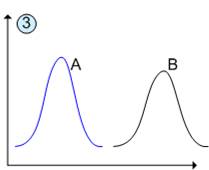
Якщо екологічні ніші організмів двох видів сильно відрізняються один від одного, то ці види, що мають одне і те ж місце проживання, не конкурують один з одним (рис.3).
Конкуренція призводить до важливих екологічних наслідків. В природі особини кожного виду одночасно піддаються міжвидовій і внутрішньовидовій конкуренції. Міжвидова конкуренція за своїми наслідками протилежна внутрішньовидовій, тому що вона звужує площу місцеперебування, кількість і якість необхідних ресурсів середовища.
Внутрішньовидова конкуренція сприяє територіальному поширенню видів, тобто розширенню просторової екологічної ніші. Кінцевий результат – співвідношення міжвидової і внутрішньовидової конкуренції. Якщо міжвидова конкуренція більша, то ареал даного виду зменшується до території з оптимальними умовами і одночасно збільшується спеціалізація виду.
Людина як біологічний вид займає свою екологічну нішу. Людина може жити в тропіках і субтропіках, на висотах до 3-3, 5 км над рівнем моря. Реально в даний час людина живе в значно більших просторах. Людина розширила вільну екологічну нішу завдяки використанню різних пристосувань: житла, одягу, вогню тощо.
|
|